
Справочник «Биофизики России»
РЕЖАБЕК Борис Георгиевич
кандидат биологических наук 03.00.03 — биофизика (бионика, 108)
Международный Экологический Фонд –Председатель правления
Северо-Кавказского Отделения
Институт Ноосферных Разработок и Исследований (ИНРИ) — Директор
Международный Университет междисциплинарных исследований —
Зав. Кафедрой нанобионики
Основные работы по биофизике и биологической
кибернетике:
Исследование динамики сверхслабых свечений возбудимых тканей (совм. с Э. М. Рудой) // Труды II-й Всесоюзной конференции по биолюминесценции, Новосибирск, 1967
Исследование динамики аутофлуоресценции возбудимых структур (совм. с В. Н. Радченко) // Труды II-й Всесоюзной конференции по биолюминесценции, Новосибирск, 1967
Принцип Бауэра и биокибернетика. // Труды I-й Северо-Кавказской конференции по кибернетике. Ростов н/Д, 1968
О некоторых микроструктурных и свободнорадикальных показателях возбуждения. // «Физико-химические основы процессов возбуждения» М, МОИП, 1970 (совм. с А. Б. Коганом и Л М. Гоголевой) с. 91—96
Самонастройка в одиночном нейроне. // Материалы IV Всесоюзной конференции по нейрокибернетике. Ростов н/Д, 1970
Самонастройка в одиночном сенсорном нейроне, как модель возможного поведения элемента самоорганизующихся нейронных ансамблей.
Автореферат дисс. на соискание учёной степени кандидата биологических наук. Ростов н/Д, 1970
О поведении механорецепторного нейрона в условиях замыкания его цепью искусственной обратной связи. ДАН СССР, 1970, т. 196, вып.4. С. 981—984
Автоколебания в системе, включающей нервную клетку в цепи искусствен- ной обратной связи. //Конференция «Колебательные процессы в биологических и химических системах». Пущино-на Оке, 1971
Биологическая кибернетика. Учебное пособие для университетов. (соавт.- А.Б.Коган, Н.П.Наумов, О. Г. Чораян). М. «Высшая школа», 1972.
Нелинейные колебания, как механизм регулирования ритмической активности нейрона (соавт.- А.Б.Коган, О. С. Васильева)
// Материалы IV Международного Конгресса по биофизике, М. 1972
Нелинейное захватывание ритма при раздражении изолированной нейрорецепторной клетки // ДАН СССР, 1973 т. 210, вып.2 (соавт.
А.Б.Коган, О. С. Васильева)
Захватывание ритма при раздражении нейрорецепторной клетки
«Биофизика», 1973, т. XVIII, №2 (соавт. А. Б. Коган, О. С. Васильева)
Устойчивое неравновесие живой материи — основа избирательной чувствительности биологических объектов к электромагнитным полям.
// «Электромагнитные поля в биосфере» М., 1985, «Наука» Т.2, с. 5—16
Общие аспекты исследования механизмов биологиче ского действия ЭМП в биологии (соавторы Н.Ф.Красногорская, Л.А.Шелепин, В. Е. Быховский).
// там же, Т.2, с. 171—178
Развитие и современное состояние проблемы биологических усилителей
// II Международный Конгресс «Слабые и сверхслабые поля и излучения в биологии и медицине». СПБ,2003
О ПОВЕДЕНИИ МЕХАНОРЕЦЕПТОРНОГО НЕЙРОНА В УСЛОВИЯХ ЗАМЫКАНИЯ ЕГО ЦЕПЬЮ ИСКУССТВЕННОЙ ОБРАТНОЙ СВЯЗИ
Доклады Академии наук СССР
1971. Том 198, №4 ФИЗИОЛОГИЯ
УДК 62—506+612.815.1
(Представлено академиком Е. М. Крепсом 21 IV 1970)
Обсуждение принципов, обеспечивающих высокую надежность биологических управляющих систем, по сравнению с техническими, приводит к представлению о бессхемной вероятностно-статистической организации структур ц. н. с.2 Роль отдельных нейронов в таких структурах фиксирована не жестко, что приводит к известной взаимозаменяемости элементов и обеспечивает надежность. Однозначность реакции системы достигается при этом за счет статистического усреднения реакций отдельных нейронов. Существует ряд экспериментальных данных (3), указывающих на то, что в структурах мозга нейроны объединяются в функциональные ансамбли, организо ванные по вероятностно-статистическому принципу.
Изучение механизмов самоорганизации нейронных ансамблей должно проводиться на базе исследования тех особенностей, которые характерны для поведения нейронов, как элементов биологических систем.
К таким свойствам в первую очередь относиться «активность», т. е. способность биологических систем к таким перестройкам структуры и функции, которые обеспечивают достижение оптимального режима функционирования уже на уровне отдельного элемента1.
Объединение таких элементов в системы более высокого порядка приводит к известным ограничениям в достижении отдельными элементами их «частных» оптимальных режимов.
Тем не менее наличие активности на уровне элементов представляется характерной и принципиально важной чертой биологических самоорганизующихся систем. В настоящей работе предпринята попытка выявить такие свойства на уровне отдельного нейрона.
Удобным объектом для исследований такого рода является нейрорецептор растяжения речного рака4, допускающий изучение поведения нервной клетки, изолированной от регуляторных ограничений со стороны организма, чего при работе с другими объектами добиться чрезвычайно трудно.
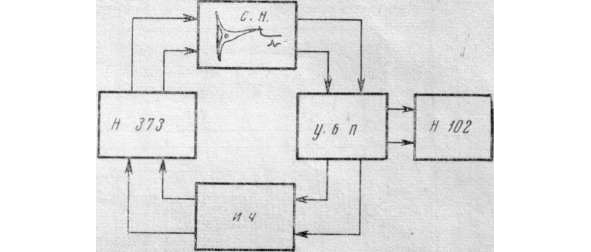
Было проведено выяснение характера поведения механорецептора в условиях замыкания его цепью искусственной обратной связи по схеме на рис.1.
Импульсная активность нейрона отводилась внеклеточно при помощи усилителя биопотенциалов УБП1—02 и регистрировалась с помощью шлейфного осциллографа Н-102.
Частота импульсов нейрона связана со степенью растяжения рецепторной мышцы (4). Один из концов этой мышцы был закреплен неподвижно, а другой захватывался с помощью зажима, который был соединен со стрелкой самопишущего вольтметра рычагом с отношением плеч 1:8.
На вход вольтметра поступал сигнал с измерителя частоты, пропорциональный частоте импульсацни нейрона.
Обратная связь, устроенная таким образом, приводила к тому, что увеличение частоты перемещало стрелку в сторону, уменьшающую растяжение мышцы, а уменьшение частоты влекло за собой увеличение растяжения мышцы.
В начале опыта стрелку фиксировали в таком положении, при котором нейрон генерировал импульсы с частотой 5—7 имп/сек.
При выключенной обратной связи препарат может работать с такой частотой много часов.
Включение обратной связи (о.с.), производимое путем освобождения стрелки, приводит к тому, что в системе возникают автоколебания: при растяжении мышцы частота возрастает, это приводит к сжатию, частота уменьшается (в силу особенностей динамических характеристик рецептора частота при быстром сжатии обычно падает до нуля), затем мышца вновь растягивается и т. д. (рис. 2а, б).
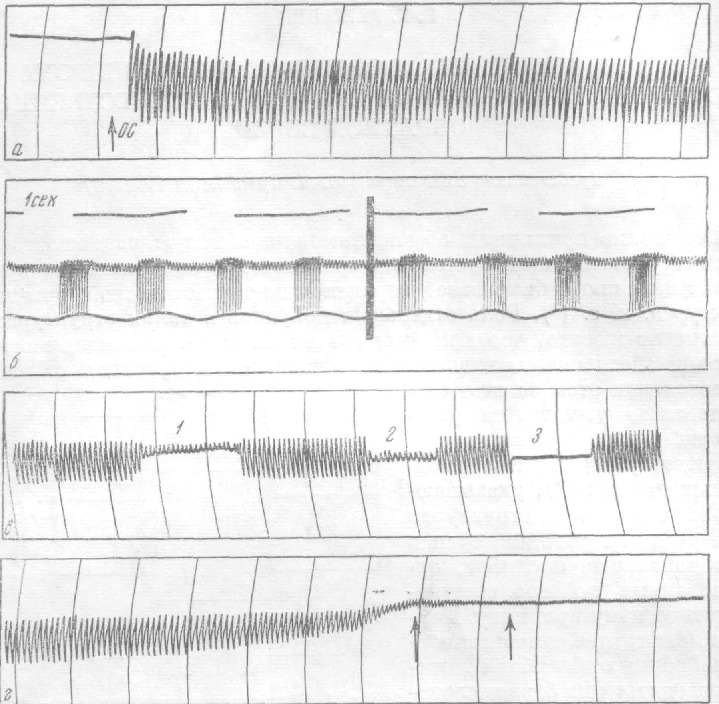
Рис. 2. Устойчивые автоколебания и п. с. р.: а — возникновение автоколебаний; о.с. — момент включения обратной связи. Запись на Н-373 в течение 100 сек. б — возникновение автоколебаний. Запись на Н-102. Нижний луч — колебания растяжения мышцы, средний — импульсная активность. в — ограничение автоколебаний: 1 — ограничение растяжения мышцы с одной стороны, 2 — с двух сторон, 3 — искусственная остановка. Устранение ограничений приводит к восстановлению амплитуды автоколебаний. Запись на Н-373. г — поиск стационарного режима (п.с.р.). Запись на Н-373. Последняя стадия процесса. Стрелки соответствуют моментам начала записей на Н-102
Если искусственно ограничить амплитуду качаний стрелки или зафиксировать ее на некоторое время, то колебания возникают вновь с прежними параметрами, как только вмешательство экспериментатора прекращается (рис.2в). В 12 опытах, проведенных при температуре 20°, наблюдалось следующее. Через 10—15 мин. амплитуда колебаний начинает уменьшаться, изменения частоты импульсов и степени растяжения мышцы прогрессивно убывают и в конце этого процесса система приходит в такое состояние, при котором не подвергается более ритмическому автораздражению и генерирует импульсы с постоянной частотой (рис. За). Этот процесс мы обозначаем как «поиск стационарного режима» (п.с.р.). Длительность стадии устойчивых автоколебаний и п. с. р. несколько варьируют для разных препаратов и сильно зависят от температуры. Стационарное состояние с частотой 3—6 нмп/сек может длиться несколько часов.
Это говорит о том, что физиологическое состояние препарата не страдает в период стадии незатухающих колебаний и п. с. р. Сам процесс п. с. р. не может быть сведен к обычной адаптации или утомлению препарата при действии длительного ритмического раздражения: специально поставленные опыты показывают, что нейрон может подвергаться внешним раздражениям в ритме автоколебаний до часа без существенного изменения характера своей ритмической активности.
В стационарном режиме частота может несколько изменяться. Вариации частоты имеют характер небольших случайных флуктуаций с возвращением к стационарному режиму
Изменения, происходящие в системе в процессе п. с. р., следует отнести на счет функциональных перестроек в нейрорецепторе. Пока нет оснований для ответа на вопрос о том, где локализуются эти изменения — в дендритном аппарате, ответственном за возникновение генераторного потенциала, или в аксонном холмике, где возникает ритмическая активность.
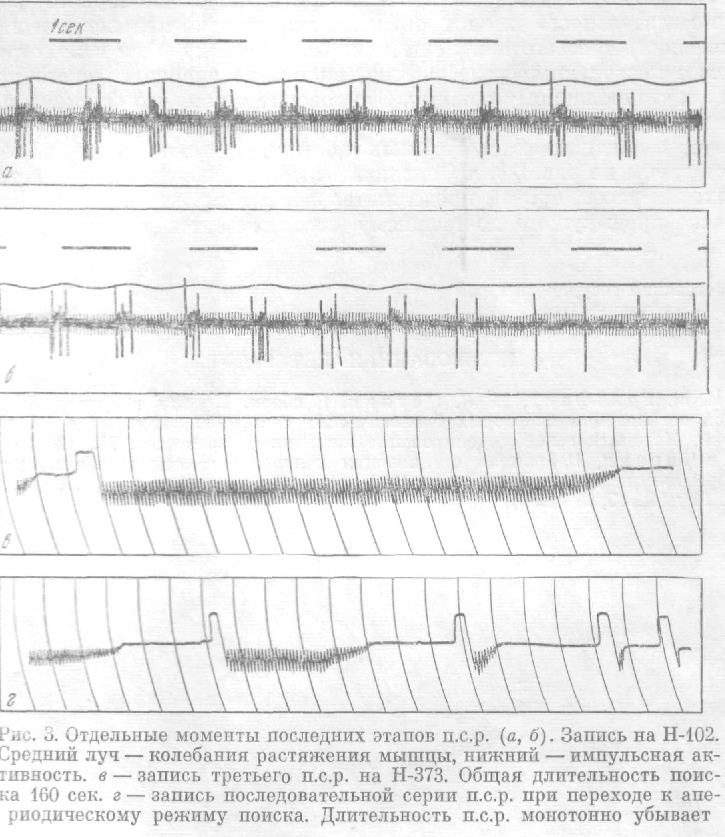
Следующий ряд наблюдений касается изменения длительности п. с. р. при повторных искусственных отклонениях от стационарного режима. Обычно длительность уже второго п. с. р. значительно меньше, чем первого. Длительность третьего п. с. р. еще меньше, а последующие п. с. р. ограничены всего несколькими колебаниями, вплоть до апериодического приближения к стационарному режиму.
Это говорит о том, что функциональные изменения, происходящие в нейрорецепторном аппарате, имеют стойкий характер и накапливаются от опыта к опыту. Примеры таких наблюдений представлены на рис. 3 в, г.
Описанные изменения сохраняются в течение нескольких часов, причем продолжительность п. с. р. через длительное время после предыдущего по-прежнему ограничивается одним-двумя колебаниями.
Поступило 16.III. 1970
ЛИТЕРАТУРА
1.И.М.Гельфанд, М.Л.Цетлин //Модели структурно- функциональной организации некоторых биологических систем, М., 1966.
2.А.Б.Коган //Математическое моделирование жизненных процессов. М., 1968, стр. 211.
3.О.Г.Чораян Нейронная организация центрального отдела зрительного анализатора лягушки. Ростов-на-Дону,1969.
4.С.Eyzaguirre,S.W.Kuffler,J.Gen.Physiol.,39,69, 1955.
О возможных внутриклеточных механизмах запоминания
Доклад в Московском Обществе Испытателей Природы (МОИП) 23 ноября 2016 года
До конца 19 века физиологи не знали, как нервные клетки связаны друг с другом. К. Гольджи считал, что между нервными клетками нет границ и нервная система представляет собой сеть. А С. Рамон-и-Кахаль считал, что нервная система состоит из отдельных нервных клеток — нейронов. Следующий шаг сделал знаменитый нейрофизиолог Чарлз Скоттт Шеррингтон, получивший Нобелевскую премию по физиологии и медицине в 1932 году «за открытия, касающиеся функций нейронов». Его учениками были такие выдающиеся нейрофизиологи, как Джон Экклз, Рагнар Гранит и Говард Флори, тоже ставшие нобелевскими лауреатами. Ч. С. Шеррингтон создал теорию синаптической связи нейронов, в 1897 году предложил понятие «синапс» и привел доказательства существования синапсов. Шеррингтон (как и Дж. Экклс) был верующим человеком и держался идеалистических взглядов на мир, его взгляды в СССР были мало известны, в основном их упоминали в связи с полемикой Ч. С. Шеррингтона и И. П. Павлова о природе сознания и теории рефлексов.
Наиболее популярной гипотезой о природе памяти в нейрофизиологии и сегодня является идея о том, что при работе синапсов их проводимость увеличивается. Поэтому многие нейрофизиологи опираются на предпо- ложение о том, что память и накопление информации связаны с изменениями в синапсах. Большинство современных нейрокибернетических моделей мозга исходит из этих представлений. В основе таких моделей лежит «нейрон Мак-Каллоха — Питтса» (1943), фактически — пороговый триггер со многими входами.
Нейрон Мак-Каллока — Питтса
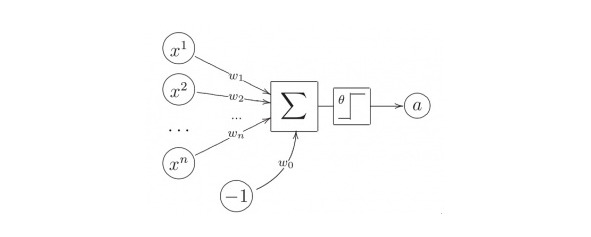
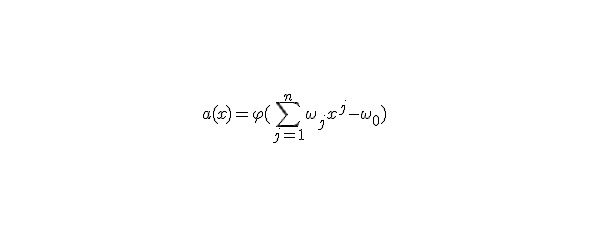
В 1943 г. Уоррен Мак-Каллок и Уолтер Питтс предложили модель нейрона, обладающую следующими свойствами:
— нейрон имеет несколько входов и один выход
— через входы (j) нейрон принимаетвходные сигналы (xj)
— входные сигналы умножаются навесовой коэффициент (wj) и суммируются
— выходной сигнал нейрона — нелинейная функция f (S),
называемая активационной. Обычно φ — это функция Хевисайда.
Было установлено, что НС могут выполнять любые логические операции и любые преобразования, реализуемые дискретными устройствами с конечной памятью.
Эта модель легла в основу теории логических сетей и конечных автоматов и активно использовалась психологами и нейрофизиологами при моделиро- вании некоторых локальных процессов нервной деятельности.
Персептрон Розенблатта
Идея Мак-Каллока — Питтса была реализована Фрэнком Розенблаттом в 1958 г. в виде компьютерной программы, а в 1960 г. в виде электронного устройства, моделирующего сетчатку человеческого глаза. Позже М. Минский и С. Пайперт в своей книге «Персептроны» строго математически доказали, что однослойные персептроны в принципе не способны решать многие простые задачи. Многие понимали, что надо усложнять структуру персептронов, и были разработаны «многослойные персептроны»
Задачи, решаемые с помощью нейросетей
— если есть математическая модель какого-то процесса, то изучая влияние входных параметров на выходные, можно решить задачу оптимизации моделируемого процесса
— если математическая модель является нестационарной, то её можно использовать для решения задач прогнозирования
— если математическая модель работает в реальном режиме времени, то результаты математического моделирования могут быть оперативно переданы оператору, управляющему объектом, или могут быть непосредственно введены в приборы, что позволяет решать задачи управления моделируемым объектом или процессом
— нейронные сети могут решать задачи распознавания и классификации образов, причем под образами понимаются зрительные изображения, символы, тексты, запахи, звуки, шумы.
Сегодня существует фактически два типа вычислительных систем для компьютеров — Фон-Неймановская и нейрокомпьютерная. Но пока что никакой из них не удалось описать реальное поведение даже простейшей нервной системы нематоды Caenorhabditis elegans!
В 60-е годы большие ученые делали многообещающие прогнозы по поводу возможностей компьютерного моделирования живых систем, и человеческого мозга в частности.
Норберт ВИНЕР в работе « Кибернетика и общество» писал:
«Индивидуальность тела есть скорее индивидуальность огня, чем индивидуальность камня, это индивидуальность формы строения, а не кусочка вещества. Эта форма может быть передана или видоизменена и скопирована, хотя в настоящее время мы лишь знаем, как скопировать ее на близком расстоянии (…)
То, что мы пока не можем телеграфировать схему человека из одного места в другое, связано, в основном, с техническими трудностями…»
Иосиф Самуилович ШКЛОВСКИЙ (в книге «Вселенная, жизнь, разум» 1975): « За прошедшие 15 лет «разум» наших электронных вычислительных машин улучшился в миллион раз… В течение нескольких следующих десятилетий следует ожидать увеличения характеристик «разума» машин еще по крайней мере в несколько десятков тысяч раз. «Разум» таких машин по основным пара- метрам будет заведомо превосходить разум человека».
Через 40 лет (в 2015 году) компьютерные мощности увеличились примерно в миллиард раз. Но где же машины, которые заведомо превосходят разум человека по основным параметрам?
Понимание ситуации было и остается фундаментально неверным!
Учет возможностей нервных клеток к поиску оптимального состояния, обучению и запоминанию результатов, открывает совершенно новые пути для понимания работы мозга и, возможно, созданию новых устройств на этой основе. В США на эти проблемы тратят не менее 100 миллионов долларов в год, но дальше идеи о «коннектоме» — составлении карты связей между нейронами) пока что не продвинулись.
Очевидно, современным создателям «Искусственного интеллекта» явно не хватает какого-то принципиально нового «измерения», нового подхода, который мог бы позволить понять принципиальную разницу между работой живого мозга и мёртвой электронной схемы. По нашему убеждению, в этой сфере могут оказаться востребованными идеи В. Я. Александрова, которые вдохновили автора на проведение экспериментов с нервной клеткой, вообще не обладающей синапсами — рецепторе растяжения речного рака.
В эксперименте на изолированном нейроне, замыкая его обратной связью по частоте, автор обнаружил способность одиночного нейрона к поисковой активности, достижению через поиск оптимального состояния и запоминания кратчайшего пути, ведущего к этому состоянию (Доклады АН СССР, 1971, т.198, №4, с. 981—984)
Было показано, что даже отдельный нейрон способен к довольно сложному поведению, обучению и запоминанию результата. Результат опыта представляет интерес как для создания более близких к оригиналу моделей мозга, так и для цитоэтологии. Владимир Яковлевич Александров (1906–1995), создатель «цитоэтологии» — науки о поведении живых клеток, писал, что у живой клетки «есть маленькая, но душа». (В.Я.Александров. «Проблема поведения на клеточном уровне.» Успехи современной биологии. 1970. Т.69, вып. 2. C. 220—248)
Ричард Фейнман (1918 — 1988), один из величайших физиков ХХ века, получивший нобелевскую премию за работы в области квантовой электродинамики, проявлял большой интерес к этой проблеме. Он писал:
«Однажды я наблюдал за парамецием и увидел что-то, что не было описано в книгах, по которым мы учились в школе, более того, в учебниках для колледжа этого тоже не было. (…) В учебниках это поведение описано как чисто механическое, подобное компьютерной программе — но выглядит оно совсем не так. (…) Оно далеко не так механистично или одномерно, как там написано.
Поведение этих простых животных следовало бы описывать правильно. Пока мы не увидим истинный масштаб поведения одноклеточного животного, мы не сможем до конца понять поведение более сложных животных».
Представляет интерес проблема передачи информации от мембраны клетки к ДНК, выяснение природы процессов, позволяющих изменять активность генов под влиянием сигналов, связанных с усилительными клеточными системами.
ПРОТОКОЛ №08/016/ПЭМ заседания секции МОИП «ПРОБЛЕМЫ ЭВОЛЮЦИИ МАТЕРИИ»
23 ноября 2016 года г. Москва
ПРИСУТСТВУЮТ: д. ч. МОИП С. В. Багоцкий, Г. Л. Ванециан, д. ч. МОИП И. А. Верещагин, И. А. Карышева, д. ч. МОИП А. П. Кугай, д. ч. МОИП Ю. М. Люляев, д. ч. МОИП К. А. Мусин, д. ч. МОИП Б. Г. Режабек, д. ч. МОИП А. Ф. Топунов, А. Л. Харитонов, К. Б. Шумаев, д. ч. МОИП Е. Д. Яхнин.
ПОВЕСТКА ДНЯ:
1. Доклад Б. Г. Режабека «О возможных внутриклеточных механизмах запоминания».
Е. Д. Яхнин: открыл заседание и предоставил слово Б. Г. Режабеку.
Б. Г. Режабек сделал доклад о своих экспериментах на изолированной нервной клетке, замкнутой цепью обратной связи, в которых было продемонстрирована способность нейрона к поиску оптимального режима, обучению и запоминанию результата. (см. выше)
В 1971 году докладчик исследовал одиночный нейрон, прикрепленный к мышце и реагирующий на её растяжение.
Он замкнул нейрон системой обратной связи, заставляющей сжиматься мышцу. Возникли автоколебания. Но появилось и нечто более интересное.
Оказалось, что характеристики пачек импульсов со временем меняются. Через полчаса нейрон входил в стабильный режим — автоколебания прекращались.
Но при повторении эксперимента автоколебания прекращались уже через 15 мин. А в третьем эксперименте колебания прекратились менее чем за минуту. При этом наблюдались изменения интервалов между импульсами — наблюдалась «поисковая активность».
Это свидетельствует о том, что нейрон способен к поисковому поведению и самообучению, причем запоминание результатов поиска не может быть связано с изменениями в синапсах, поскольку таковые отсутствуют. Этот результат находится в соответствии с представлениями В. Я. Александрова, который выдвинул идею о «поведении клеток». В 1970 году он опубликовал в «Успехах современной биологии» статью «Цитоэтология». Эти идеи поддерживал и Ю. А. Лабас.
В 1975 году И. С. Шкловский писал «Разум вычислительных систем увеличился с 1960 года в миллионы раз. И скоро он превзойдет человеческий мозг». Однако это предсказание не оправдалось. Не реализовалась и компьютерная модель мозга.
Физик Р. Фейнман считал, что инфузории ведут себя не так, как их модели.
Дж. Экклз говорил о душе. По его мнению, главная задача сегодня заключается в том, чтобы спасти человека от материалистической деградации.
А.Л.Харитонов: Каковы отношения Вашего эксперимента к гомеостатике?
Б. Г. Режабек: Это не совсем гомеостатика. Опыты свидетельствуют о том, что у клетки есть свои потребности.
А. Ф. Топунов: Какие ещё были работы на эту тему? С момента Вашей публикации прошло 45 лет!
Б. Г. Режабек: В 70-е годы моя лаборатория в Ростовском университете была разрушена, и я не имел возможности продолжать эксперименты. Вы не хотели бы их продолжить?
А. Ф. Топунов: Я занят другими проблемами. Вы пытались публиковаться на Западе?
Б. Г. Режабек: Нет.
А. П. Кугай: Сколько времени сохранялась память о предыдущих экспериментах.
Б. Г. Режабек: Неизвестно. Нейрон гибнет через двое суток.
А.П.Кугай: Не связаны ли Ваши результаты с утомлением нейрона?
Б.Г.Режабек: В экспериментах без обратной связи утомления не было.
Клетка влияет на окружающую среду и находит для себя оптимальное состояние.
К.А.Мусин: А почему Вы называете это поиском?
Б.Г.Режабек: Потому что эти изменения случайны.
Ю.М.Люляев: Сколь правомерны аналогии между клеткой и человеческим обществом?
Б.Г.Режабек: Столь же правомерны, как понятие «поведение».
А.Л.Харитонов: Свойствами, которые Вы обнаружили, в принципе обладает любая большая макромолекула.
Б.Г.Режабек: Наверное, да.
А.Л.Харитонов: Нужно исследовать разные системы. Любая система ищет устойчивость во внутренней системе координат.
К.А.Мусин: Как можно использовать теорию конечных автоматов для анализа клеток?
Б. Г. Режабек: Не знаю.
Ю.М.Люляев: Существует принципиальная разница между сознанием и поведением клеток. Человек познает мир душой. Человеческое сознание принципиально общественно.
Б.Г.Режабек: Слово «поведение» очень важное. У собаки есть поведение, а у пули — нет.
Ю. М. Люляев: Поведение есть там, где возможен выбор.
И.А.Карышева: В Индии жил великий мыслитель Шри Ауробиндо, в работах которого говорится о «разуме клеток».
Б. Г. Режабек: Шри Ауробиндо — это не наука, а метафизика. Но индийский биофизик Джагадиш Чандра Бос имел сходные представления.
В беседе с С. В. Багоцким мы пришли к выводу о том, что МОИП мог бы представить нашу работу на соискание Нобелевской премии. Я делал доклад в МОИП о Нобелевских премиях, не выданных русским ученым, и говорил о том, что в ряде случаев (А. М. Оловников, Ю. А. Лабас) это произошло из-за того, что Европа мало читает нашу научную литературу, а мы не думаем о наших приоритетах и не представляем работ наших учёных в Нобелевский комитет.
А.Ф.Топунов: Экспериментальные данные докладчика очень интересны, но объяснение без дальнейших экспериментов — это метафизика. Нужно искать гранты на продолжение работы. Кто хочет — ищет возможности, кто не хочет — ищет причины. У эритроцитов есть метаболическая память.
Нужны серьезные исследования. Вопрос о выдвижении на Нобелевскую премию наше собрание решать не может, но можно поставить его на общей Конференции МОИП.
Е. Д. Яхнин: Ваши эксперименты очень интересны и нуждаются в развитии. Но я не могу одобрить Ваше стремление всюду искать душу. Душа — это не объяснение.
Председательствующий поблагодарил докладчиков и участников заседания за интересную дискуссию и объявил заседание закрытым.
Председатель секции Проблем
Эволюции материи МОИП Е.Д.Яхнин
Ученый секретарь секции А. К. Погорелов
Ученый секретарь МОИП С. В. Багоцкий
ЗАХВАТЫВАНИЕ РИТМА ПРИ РАЗДРАЖЕНИИ
НЕЙРОРЕЦЕПТОРНОЙ КЛЕТКИ РЕЧНОГО РАКА
БИОФИЗИКА Том XVIII, вып. 21973
А. Б. Коган, Б. Г. Режабек, О. С. Васильева
Ростовский государственный университет
При локальном раздражении синусоидальным током через микроэлектрод быстро адаптирующегося и медленно адаптирующегося нейронов обнаружено захватывание частот с отношением 1: 1 в области 2—4 и 19— 23 гц, а также с отношением 2:1 в области 5—7 и 24 — 27 гц. Увеличение амплитуды стимула приводит к расширению полос захвата. Обсуждается связь наблюдаемых эффектов с проблемой самоорганизации нейронных сетей.
Проблема самоорганизации нейронных структур требует выяснения двух основных вопросов:
1) какие свойства элементов систем существенны для самоорганизации, 2) какими должны быть законы взаимодействия элементов, позволяющие реализовать самоорганизацию.
То, что элементы самоорганизующихся систем должны обладать определенной спецификой, неоднократно подчеркивали как математики [1—3], так и биологи [4—8]. Поведение элементов биосистем в свою очередь определяется спецификой физико-химического состояния живого вещества, находящегося в состоянии «устойчивого неравновесия» [6], которое, с современной точки зрения, можно интерпретировать, как результат взаимодействия многоуровневых внутри- клеточных нелинейных осцилляторов как кинетической, так и молекулярно-структурной природы.
По-видимому, механизм взаимодействия нелинейных осцилляторов является одним из механизмов самоорганизации [9, 10].
В особенности существенным этот механизм может считаться при рассмотрении внутриклеточной самоорганизации как механизма синхронизации пространственно разобщенных процессов в клетке и взаимодействия нейронов, каждый из которых непрерывно подвергается воздействию ритмических импульсных потоков.
Хотя этот вопрос несомненно важен, в современных моделях нейронов, к сожалению, не представлены ни способность нейрона к целесообразному приспособлению, ни возможность наличия эндогенных ритмов, свойст- венных данному нейрону и позволяющих ему избирательно реагировать на входной импульсный поток. Наличие таких ритмов обсуждалось в ряде физиологических работ в связи спроблемой «пейсмекеров» [11].
В настоящей работе представлены эксперименталь- ные данные, свидетельствующие о наличии таких ритмов в изолированной нейрорецепторной клетке речного рака.
Методика
Потенциалы действия (ПД) отводились внеклеточно от нервного ствола с помощью серебряного электрода диаметром 0,3 мм. Импульсы усиливались УБП1—02 и регистрировались с помощью шлейфного осциллографа Н-102.
Раздражение производилось стеклянными микроэлек- тродами, заполненными 3М раствором КСl. Диаметр кончика измерялся под микроскопом МБР-1 и составлял 2—5мк, сопротивление 1—5 Мом.
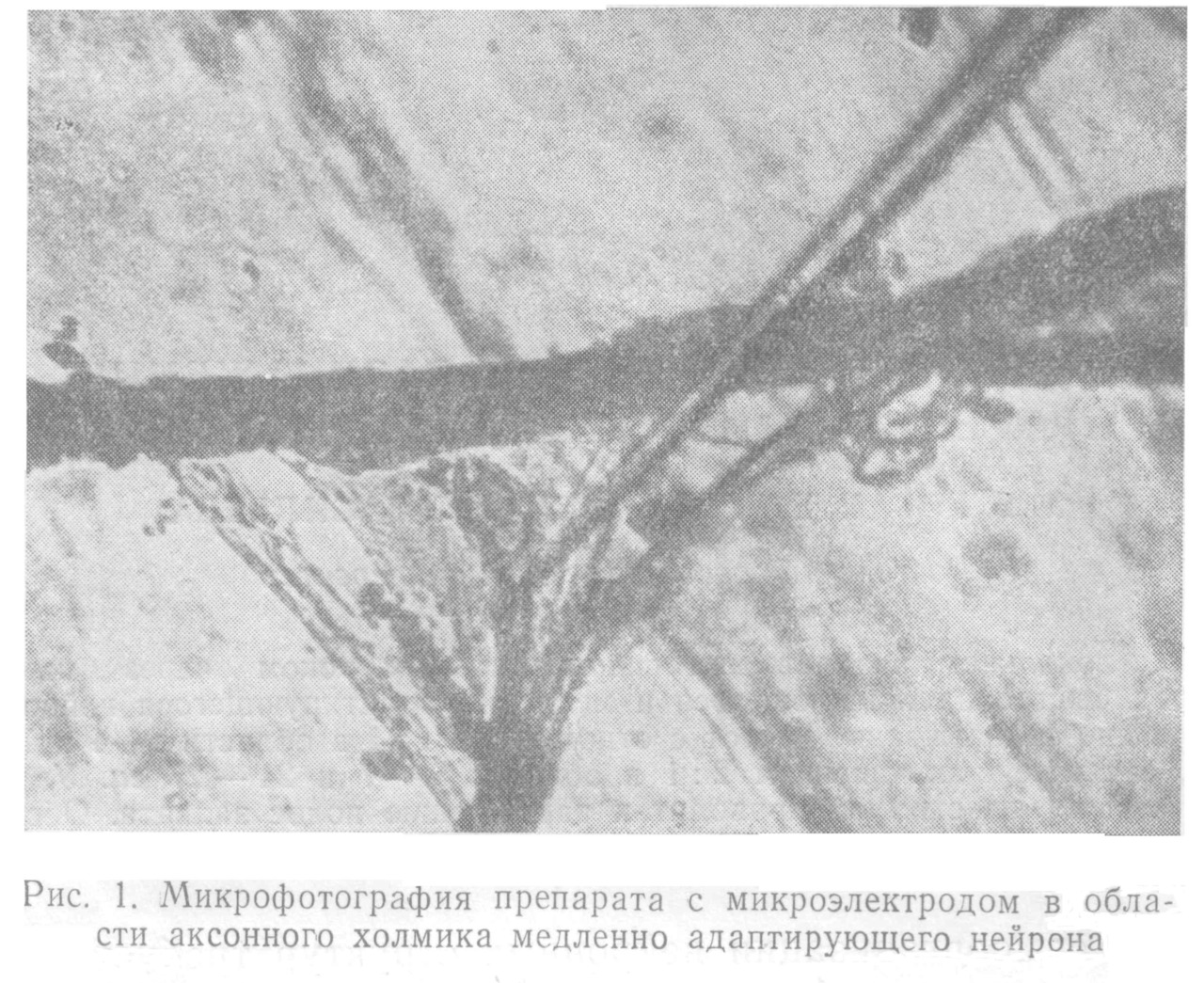
Микроэлектрод подводили с помощью микроманипулятора ММ-1 к избранной точке нейрона под контролем микроскопа МБР-1 (рис.1). В качестве источника раздражающего тока использовался генератор Г3—47.
Результаты экспериментов
— Реакция быстроадаптирующегося нейрона.
Реакция быстроадаптирующегося (БА) нейрона на механичеcкое (адекватное) раздражение достаточно подробно изучена многими авторами [12, 13].
При электрическом раздражении наиболее эффективным является положение микроэлектрода в области аксонного холмика (АХ), при этом возбуждение происходит, когда на микроэлектроде отрицательное напря- жение. Порог раздражения в этом случае минимален. Располагая микроэлектрод вблизи сомы или удаляясь от АХ по волокну, можно наблюдать изменение порога и даже инверсию знака раздражающего потенциала.
При условии одинаковой реакции на толчок постоянного тока, которой можно добиться, расположив микроэлектрод при подготовке к опыту соответствующим образом, реакции различных препаратов на ритмическое раздражение совпадают с точностью до 10%.
Такое раздражение не вызывает импульсной активности нейрона, пока fc <1,5 гц.
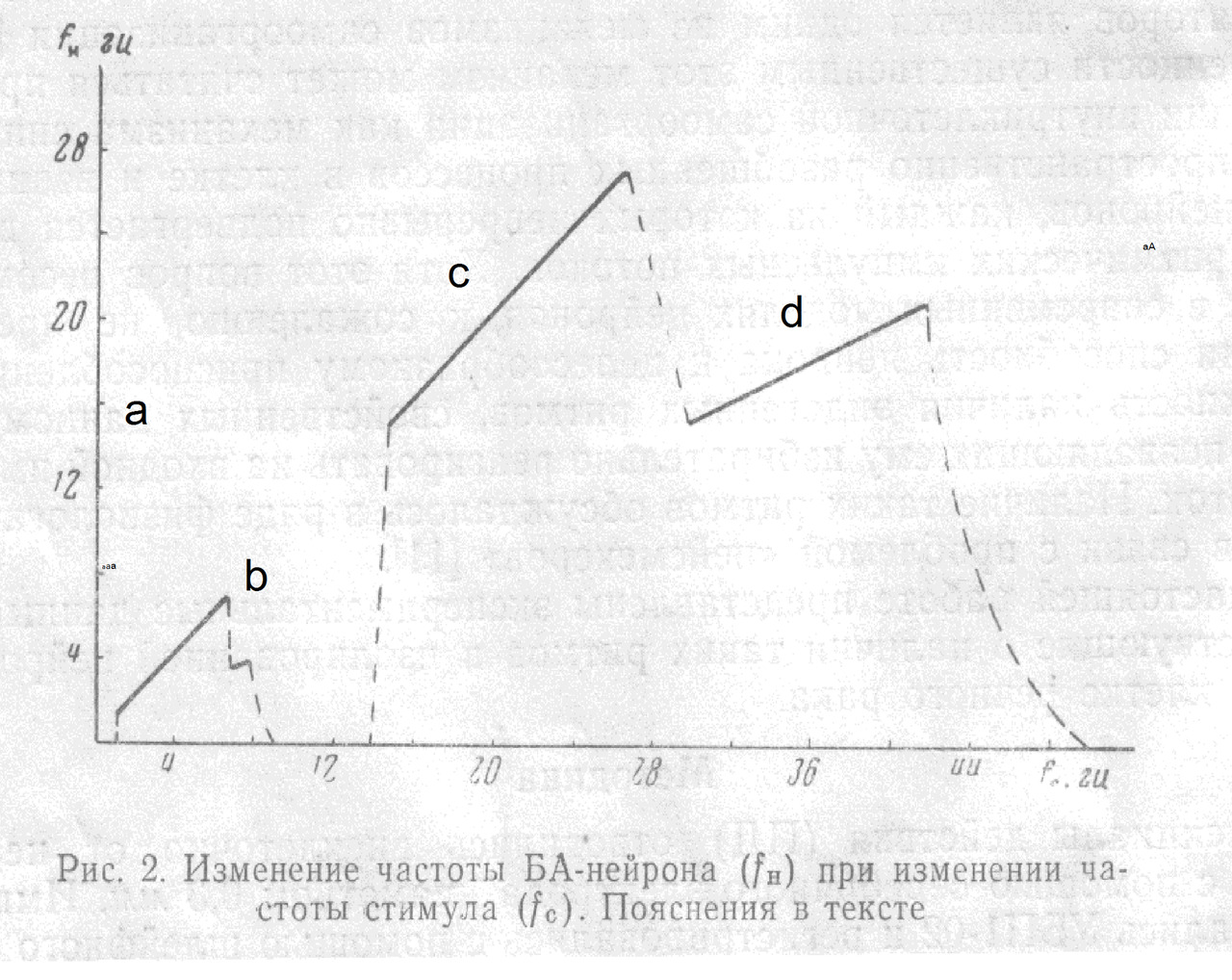
На рис. 2 показано изменение частоты БА-нейрона (fH) при изменении частоты стимула (fc); амплитуда раздражения 11 в. Микроэлектрод расположен вблизи АХ.
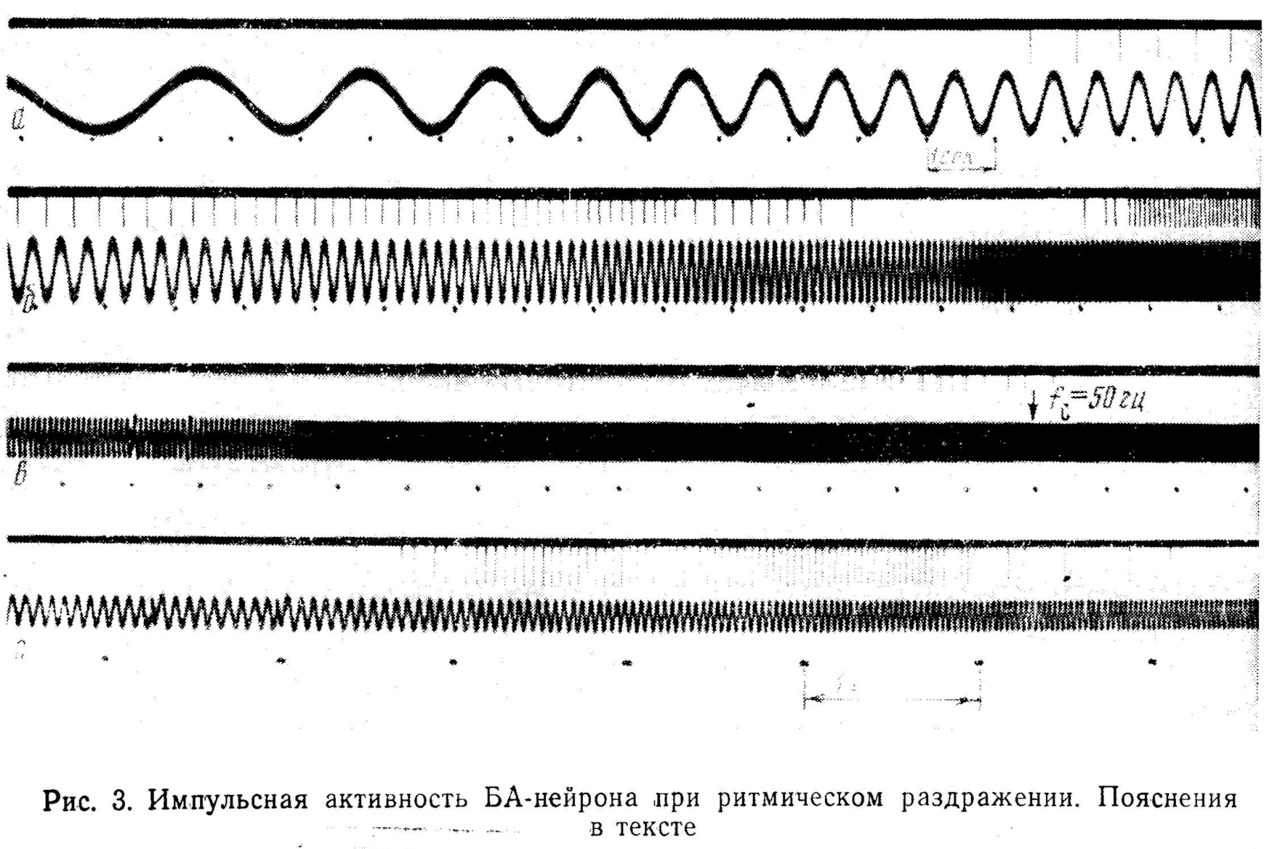
При достижении этой частоты наблюдается резкий переход к режиму, при котором нейрон работает с частотой, совпадающей с fc (рис. 3,а). Диапазон захватывания частоты (ЗЧ) зависит от амплитуды стимула.
В данном опыте ЗЧ скачком прекращается при fc = 7 гц. При изменении fс от 7 до 8 гц наблюдается ЗЧ с делением в отношении 2:1, при частотах, незначительно превышающих 8 гц, нейрон реагирует на каждый третий или четвертый стимул и затем реакция вообще исчезает вплоть до частоты 13 гц (рис. 3,б). От 13 до 14 гц реакция является вероятностной, т. е. однозначного соответствия между fс и fH установить не удается и при повторных стимуляциях реакция в этом диапазоне оказывается каждый раз иной. С 14 гц начинается ЗЧ вплоть до 27 гц. В диапазоне от 27 до 30 гц наблюдается переходной режим реакции, при котором можно обнаружить отклик нейрона, соответствующий либо каждому периоду стимула, либо с выпадением одного периода. С 30 до 42 гц наблюдается ЗЧ с делением частоты 2:1, и, наконец, при более высоких частотах реакция вновь становится вероятностной.
При дальнейшем увеличении частоты стимуляции вплоть до 20 000 гц импульсная реакция нейрона отсутствует. На рис. 3, в представлен ход процесса от 15 до 60 гц. Если изменять амплитуду стимула, общий ход реакции закономерно изменяется.
Результаты можно объединить с помощью рис.4. Линия а представляет собой геометрическое место точек, соответствующих началу первой полосы ЗЧ с отношением частот fc: fa равным 1:1. Линия б соответ ствует переходу в вероятностный режим, в — началу первой полосы ЗЧ с отношением частот 2:1, г — ее окончанию и д — исчезновению импульсной реакции. Линия е соответствует началу новой полосы ЗЧ.
Реакция становится детерминированной (отношение частот 1:1), начиная с линии ж, и остается таковой вплоть до линии з. Между з и и реакция недетерминированна, между и и к захватывание происходит с делением частоты 2:1. В области между к и л реакция опять становится вероятностной, и, наконец, линия л соответствует оконча- тельному исчезновению импульсной реакции нейрона вплоть до fс= 20 000 гц. Таким образом, можно наб- людать три четко выраженных полосы ЗЧ (а-б, ж — з, и -к) и одну (в — г) менее выраженную.

Эти эффекты мы наблюдали на 30 препаратах. На одном и том же препарате измерения можно проводить многократно (10 и более раз) без заметных изменений в результатах. Для различных препаратов соответствующие частоты несколько отличаются вследствие небольших различий в положении микроэлектрода, а также в зависимости от физиологического состояния препарата.
Наиболее стабильна область второго ЗЧ с отноше- нием частот 1:1 (ж-з), в то время как предыдущие области захватывания на некоторых препаратах отсутствовали. Иногда отсутствовала и область и-к (рис.3, г).Если кончик микроэлектрода расположить вдали от АХ, общий ход кривой fн = φ (fс) изменяется.
В области низких частот в этих опытах до того, как начинается захватывание 1:1, можно наблюдать пачечную активность. Число импульсов в пачке с увеличением fc уменьшается, и пачка вырождается в одиночные импульсы, затем происходит деление частоты в 2, 3, … п раз и где-то на 10—15 гц импульсная активность исчезает. Приведенные результаты относятся к опытам, в которых частота стимула линейно нарастала со временем.
В следующей серии опытов исследовалась реакция БА-нейрона на включение стимула фиксированной частоты (fc).
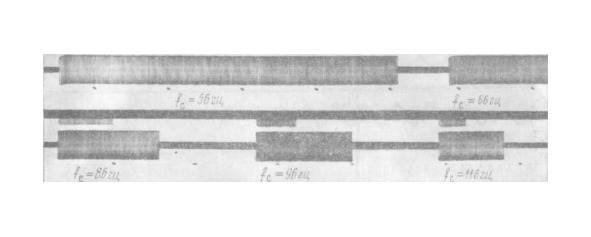
Если fc находится за пределами зон ЗЧ, то БА-нейрон некоторое время τ, зависящее от fcи амплитуды стимула, работает с частотой fн= fc, затем адаптируется и прекращает импульсную активность (рис. 5).
При fc, принадлежащей области ЗЧ τ = ∞ в этих условиях нейрон не проявляет адаптации.
Зависимости τ от fc для одного из препаратов (два первых диапазона не показаны) представлены на рис. 6.
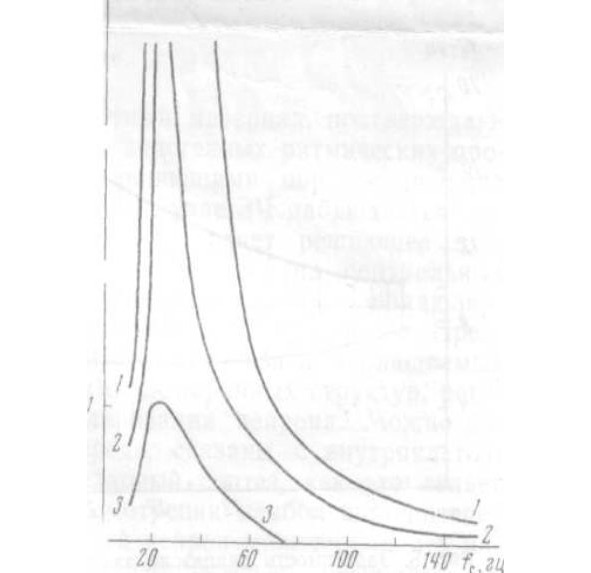
От препарата к препарату эффект варьирует незначительно.
— Реакция медленно адаптирующегося нейрона.
Реакция МА-нейрона качественно во многом сходна с реакцией БА. Основное отличие связано с тем, что МА-нейрон при отсутствии раздражения может генерировать импульсы с собственной фоновой частотой (2—30 гц), определяемой степенью приложенного растяжения.
При включении ритмического раздражения, начиная от 0,1гц, частота нейрона модулируется изменениями стимула (рис.7,а), затем появляются пачки импульсов (рис.7,б и в), дальнейшее увеличение fс вызывает ЗЧ с отношением 1:1 (рис.7, г).
Бесплатный фрагмент закончился.
Купите книгу, чтобы продолжить чтение.