
Московское общество испытателей природы при
Московском государственном университете им. М. В. Ломоносова
ДОКЛАДЫ МОИП. Секция Геронтологии. Т. 67
(к 25-летию Института биохимической физики РАН им. акад. Н. М. Эмануэля)

Издание выпускается ежегодно.
Издание рецензируемое, представлено в РИНЦ.
УДК 613.98; 612.67/.68
В сборнике представлены научные статьи авторов на основе
докладов, прочитанных на заседаниях Секции геронтологии
Московского общества испытателей природы за 2019—2020 гг.
Работы посвящены теоретическим, экспериментальным и клиническим аспектам старения.
Редакционная коллегия:
В.М.Новоселов, председатель секции, ответственный редактор;
В. И. Донцов, д.м.н., научный редактор;
В.Н.Крутько, к.б.н., д.т.н., профессор, редактор;
В подготовке издания принимали участие:
— НП «Национальный геронтологический центр»;
— Лаб. системного анализа и информационных технологий в меди-цине и экологии Федерального исследовательского центра «Инфор-матика и управление» РАН (ФИЦ ИУ РАН).
Электронный адрес редакции:
Анатолий Иванович Деев (20 мая 1948 — 3 июля 2020)

ДЕЕВ Анатолий Иванович, кандидат биологических наук, доцент, Российский национальный исследовательский медицинский университет им. Н. И. Пирогова (Москва), кафедра общей и медицинской биофизики.
Научные интересы: медицинская и биологическая физика, механизмы старения, старение кожи и эстетическая медицина, нейроспецифические белки и сосудистая патология мозга, свободные радикалы и старение. Публикации: https://elibrary.ru/author_items.asp.
Научное сообщество геронтологов Москвы и Российской Федерации с глубоким прискорбием извещает, что после непродолжительной болезни ушел из жизни Анатолий Иванович Деев, председатель Московского отделения Геронтологического общества при РАН, которое он возглавлял 20 лет.
Он является автором ряда книг и множества научных статей — на русском, английском и китайском языках.
До последнего времени он был сопредседателем секции по фундаментальной геронтологии на ежегодных Международных конференциях. Анатолий Иванович был мечтателем в науке.
Его мечта, чтобы самые обычные люди жили дольше, в здоровье и радостях. Он неустанно работал, совершенствуя познания всего человечества об этом пути.
Сегодня, когда он ушел из жизни, мы не только будем вспоминать добрым словом этого мягкого, тактичного, интеллигентного человека, мы будем надеяться, что его душа с нами. Как вместе с нами и души великих геронтологов прошлого. Мы едины с ними, мы звено в цепочке между прошлым и будущим.
Светлая память о нашем коллеге, талантливом ученом, добром,
разностороннем и отзывчивом человеке, навсегда останется с нами.
Лучшая память о нем — это продолжение его дела.
ИСТОРИЯ ГЕРОНТОЛОГИИ
ДОКАЗАТЕЛЬНАЯ ГЕРОНТОЛОГИЯ: ВЧЕРА, СЕГОДНЯ, ЗАВТРА (к 25-летию Института биохимической физики РАН им. акад. Н. М. Эмануэля)
В. Б. Мамаев
Институт биохимической физики РАН им. акад. Н. М. Эмануэля
Показана история развития геронтологии в России на примере Института биохимической физика РАН им. акад. Н. М. Эммануэля: теоретические и экспериментальные разработки по биохимии и биофизической химии старения, начатые академиком Н. М. Эммануэлем, преимущественно в области свободно-радикальной теории старения и влияния антиоксидантов на процессы старения и продолжительность жизни экспериментальных животных.
Ключевые слова: старение, история старения, теории старения.
Начало Начал.
Издавна люди мечтали продлить молодую жизнь. Но переход от мечтаний к реализации произошел только в начале ΧΧ века, когда И. И. Мечников предложил научный подход к лечению преждевременного старения [65]. Он впервые предложил называть эту научную деятельность «геронтологией». Попытки обойтись без научного фундамента при решении этой проблемы кончались или неудачей, или трагедией. Последнее обстоятельство значительно тормозило исследования старения. Для научного решения проблемы старения необходимо было решить сразу, как минимум, две задачи:
— найти метод замедления скорости старения, соответствующий предлагаемым теоретическим предположениям,
— количественно оценить величину положительного эффекта.
Важно подчеркнуть, что в те времена, и даже в настоящее время, единодушного метода определения/измерения старения не было. Кроме того не было ВАКовской специальности «геронтология», как и научного журнала по геронтологии, а также геронтологической лаборатории в АН СССР. «Медицинский институт был, а науки (в понимании сотрудников АН СССР) не было».
Несмотря на эти сложности и негативное отношение научной общественности Академии Наук СССР к геронтологии, в середине 60-х годов прошлого столетия основатель Института биохимической физики РАН академик Н. М. Эмануэль начал научные геронтологические работы в рамках своего отдела «Кинетики химических и биологических процессов» Института химической физики АН СССР.
Очевидно, первый шаг в сторону геронтологии Николай Маркович сделал значительно раньше, в 1940 году, когда стал изучать «медленный вырожденный взрыв» при горении сероводорода в кислороде.
Он показал, что, как и при быстрых реакциях горения, медленные реакции горения протекают через промежуточные вещества свободно-радикальной природы (моно оксид серы) [9, 10] и именно это промежуточное вещество играет решающую роль в механизме цепного окисления.
Важным этапом в исследованиях была идея Н.М. относительно новых принципов управления сложными стадийными процессами, при которых условия проведения реакций (в том числе добавки ингибиторов) не задаются раз и навсегда в начальный момент, а рационально изменяются по ходу процесса [11]. В короткий срок была создана и экспериментально обоснована цепная теория жидкофазного окисления органических веществ.
В дальнейшем проблема торможения и подавления нежелательных окислительно-деструктивных процессов становится одной из основных в экспериментальных и теоретических исследованиях Н.М.
Эмануэлем и сотрудниками было открыто явление избирательного ингибирования многоцентровых цепных процессов некоторыми ингибиторами (например, нитроксильными радикалами), взаимодействующими с одним из активных центров цепной реакции и не влияющими на развитие основной цепи окисления. Выполнен теоретический анализ явления и раскрыт его механизм.
Большой научный и практический интерес представляет исследование явления синергизма в смесях ингибиторов. В работах Николая Марковича и сотрудников было установлено неизвестное ранее явление радикального взаимодействия ингибиторов в процессах окисления органических веществ, заключающееся в регенерации более эффективного ингибитора вследствие переноса атома водорода к его радикалу от молекулы менее эффективного ингибитора и лежащее в основе усиления суммарного тормозящего действия ингибиторов при стабилизации органических материалов смесями ингибиторов.
Очень важным этапом было исследование механизмов лучевого поражения и применение синтетических ингибиторов радикальных процессов. Н. М. Эмануэлем с сотрудниками установлена природа первичных свободно-радикальных молекулярных нарушений, возникающих в тканях животных под действием гамма-радиации, различия в поражении ДНК и мембран в тканях с разной радиочувствительностью. Исследован молекулярный механизм пост-радиационных изменений в биоэнергетике и биосинтетических процессах в тканях органов животных в ходе лучевой болезни.
При бурном обсуждении этих принципиально новых биохимических механизмов родилось понятие «антиоксидант». (Антиоксидант — это вещество, обладающее свойствами ингибитора свободно-радикальных реакций и уравновешивающее взрывной характер цепных свободно-радикальных реакций в организме.)
Убедительно показана на молекулярном, клеточном и организменном уровне возможность торможения развития лучевых повреждений с помощью малотоксичных ингибиторов свободно-радикальных реакций — антиоксидантов. Они проявляют защитное действие в случае лучевой болезни и болезни, вызванной введением радио-миметиков. Установлена важная роль природных биоантиоксидантов, содержащихся в мембранах клеток.
Широкий спектр и особенности биологического действия ингибиторов радикальных процессов сделали необходимым выяснения молекулярных механизмов их действия.
Молекулярные механизмы действия ингибиторов обусловлены особенностями их химического строения, и в первую очередь наличием окси- или амино-ароматической группировки. Так, для соединений фенольного ряда характерна способность к различным реакциям с отрывом атома водорода от гидроксильной группы и промежуточным образованием свободных радикалов. Кинетические исследования реакций с участием ферментов гликолиза или РНК-полимеразы позволили установить, что их торможение обусловлено свободно-радикальными продуктами авто-окисления ингибиторов. Такие продукты окисляют SH-группы ферментов с образованием дисульфидных связей, ингибируя их активность.
Исследование старения экспериментальных животных
Изучение свойств антиоксидантов-радиопротекторов позволили плавно перейти к замедлению старения организма и исследованию механизмов. В 1968 году, комментируя статью в газете «Санди Таймс» об опытах Хармана, лежащих точно в той же научной области, Н. М. писал: «Результаты, полученные доктором Харманом, следует рассматривать как вполне достоверные. При этом следует отметить, что использование для подавления вышеупомянутых патологических процессов вещества, способные „захватывать“ свободные радикалы (так называемые ингибиторы радикальных процессов), было сделано впервые советскими учеными. Однако, по нашему мнению, в дальнейшем в биологии и в медицине целесообразно применять не ионол как таковой, а различные, разумно синтезированные малотоксичные Ингибиторы Радикальных Процессов» [18].
Существенной проверкой правильности новых выбранных направлений исследования старения были выступления на ΙΧ Международном конгрессе геронтологов в 1972 году в Киеве [19, 55]. Уместно заметить, что к этому времени уже были опубликованы как статьи Хармана [84], где было показано увеличение только средней продолжительности мышей на 44,6%, так и его критиков [92]. Хармана на конгрессе не было, но вопросы к его работе витали в воздухе. Может быть, поэтому пристальное внимание на конгрессе привлек доклад Н. М. Эмануэля [19] по замедлению старения мышей антиоксидантами. Увеличение средней продолжительности жизни на 25,3% и максимальной продолжительности жизни на 55,8%, полученные в ИХФ АН СССР [41] на основе хорошо обоснованного экспериментально-теоретического фундамента, в тот момент склонили чашу весов в пользу свободно-радикальной теории.
Кинетические закономерности выживаемости и определение скорости старения в эксперименте
Под процессом старения организма, в том числе и человека, мы [50] понимаем биологический процесс, который проявляется (а мы его измеряем) на популяционном уровне. Это — универсальный, эндогенный, разрушительный биологический процесс, который приводит к увеличению вероятности смерти с возрастом. Если начало старения (увеличения вероятности смерти с возрастом) приходится на возрастной интервал 10—15 лет, то конец в возрастной группе 95+. Это означает, что процесс старения включает большую часть жизненного пути организма. Т.е. процесс старения включает: начало старения (эндогенные причины старения); возрастные патогенные процессы (предболезнь); саногенные (защитные) процессы; возрастное разрушение организма, сопровождаемое патологическими процессами (т.е. возрастными болезнями); полное уничтожение данной когорты ровесников. В литературе часто понятие старения трактуют более узко, причем каждый на свой лад. Одни старение воспринимают, как предболезнь (патогенный процесс).
Т.е. естественные возрастные изменения организма, которые приводят или сопровождают развитие болезни. Другие наоборот считают старение последним этапом жизни, когда организм превращается в дряхлое создание перед концом жизни. Определение старения, которое обосновал Б. Стрелер в 1960 году [101] (в русском переводе в 1964 году [73]), не противоречит здравому смыслу и, самое главное, дает геронтологам количественный измеритель старения (возрастное увеличение вероятности смерти, т.е. возрастную смертность).
В те времена (60 — 70-ые годы) даже в киевском институте геронтологии никто не строил кривые смертности (ни мышей, ни людей). Они изучали возрастные физиологические механизмы старения, не измеряя старение. Тогда как, для количественной оценки скорости старения мышей обычно анализировали картину смертности популяции. Так как средняя продолжительность жизни существенно зависит от факторов окружающей среды и питания, то это приводило к значительной разнице показателей выживаемости, получаемых в разных лабораториях для животных одних и тех же инбредных линий.
Для отработки метода измерения скорости старения были изучены кинетические закономерности выживаемости линейных мышей CBA, C3HA, (C57BLCBA) F1 гибридов первого поколения и (SHK) нелинейных лабораторных мышей [61]. Опыты проводились с животными 2-месячного возраста (самки), полученными из Центрального питомника лабораторных животных МЗ СССР отделения «Крюково». Мыши SHK в возрасте 7,5–8 месяцев были взяты из того же источника. Условия содержания животных соответствовали требованиям, предъявляемым к длительным экспериментам на выживаемость.
Важнейшей характеристикой всякого процесса, протекающего во времени, является его скорость. Для измерения скорости старения в экспериментальной геронтологии нет единого принятого метода, большинство исследователей пользуются для этих целей статистическими показателями, такими, как средняя и максимальная продолжительность жизни, средняя продолжительность предстоящей жизни и выживаемость. Кривые выживаемости дают наглядное представление об изменении численности популяции, но благодаря их сложности (S-образные кривые, рис.1) определение скорости смертности, величина которой меняется с возрастом, требует дополнительной математической обработки. Обработка кривых подобного типа заключается в подборе эмпирических зависимостей вида: x = A / (a + e a +b t) («логистические» или «автокаталитические» функции). Применение для обработки этих данных «пробит» метод позволяет представить кривую выживаемости в виде прямой, отсекающей на оси времени отрезок, равный возрасту при 50%-ной смертности («медиана»); из наклона прямой может быть определена средняя скорость смертности в %, т.е. этот метод позволяет определить величины, имеющие вполне определенный биологический смысл [74].
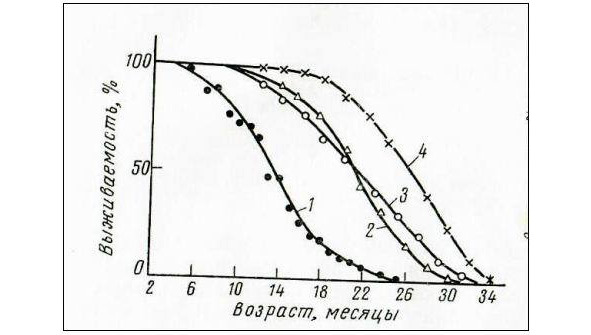
На рис. 2 представлено такое преобразование кривых выживаемости. При вычислении скорости смертности следует принимать во внимание неравномерность шкалы «пробит», благодаря чему сравнимые данные могут быть получены только с определенных участков прямой.
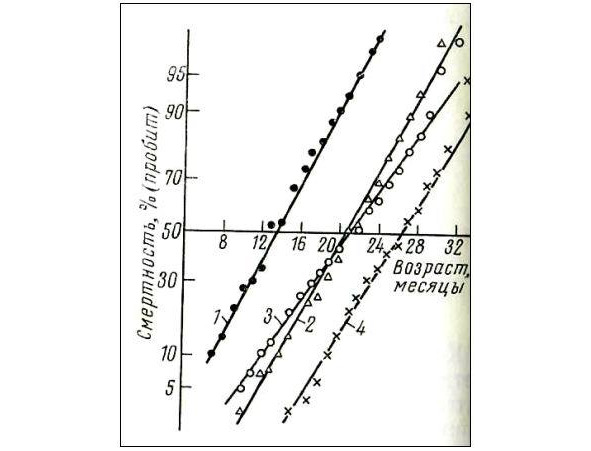
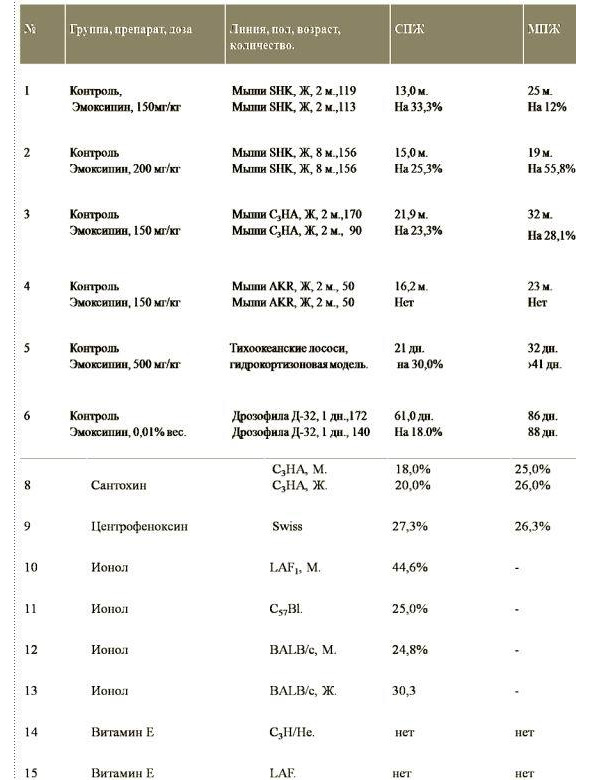
1 — SHK нелинейных лабораторных мышей, 2 — CBA, 3 — C3HA, 4 — (C57BLCBA) F1 гибридов первого поколения.
Если, как в случае прямых, изображенных на рис. 2, линейная аппроксимация удовлетворительна в интервале от 10 до 90% смертности, то можно использовать весь этот участок и вычислить среднюю скорость смертности (таблица 1), которая составляет для мышей SHK нелинейных лабораторных мышей = 6,2% в месяц; CBA = 6,2% в месяц; C3HA = 4,7% в месяц, (C57BLCBA) F1 гибридов первого поколения = 5,3% в месяц.
Известно, что гибриды первого поколения обычно жизнеспособнее животных родительских линий. Это наблюдение подтвердилось и в данной работе. Величины показателей выживаемости (C57BLCBA) F1 близки к данным, опубликованным Смитом и соавторами [99]. Величины средней продолжительности жизни мышей CBA, определенные другими авторами, значительно различаются: 27, 2; 17,5; 21,2 месяца [20, 102, 99]. Преимущества этого способа обработки экспериментальных данных особенно наглядны в тех случаях, когда посредством какого-либо воздействия (облучение, голодание, стресс, применение геропротекторов и т.п.) в ходе опыта изменяется скорость старения.
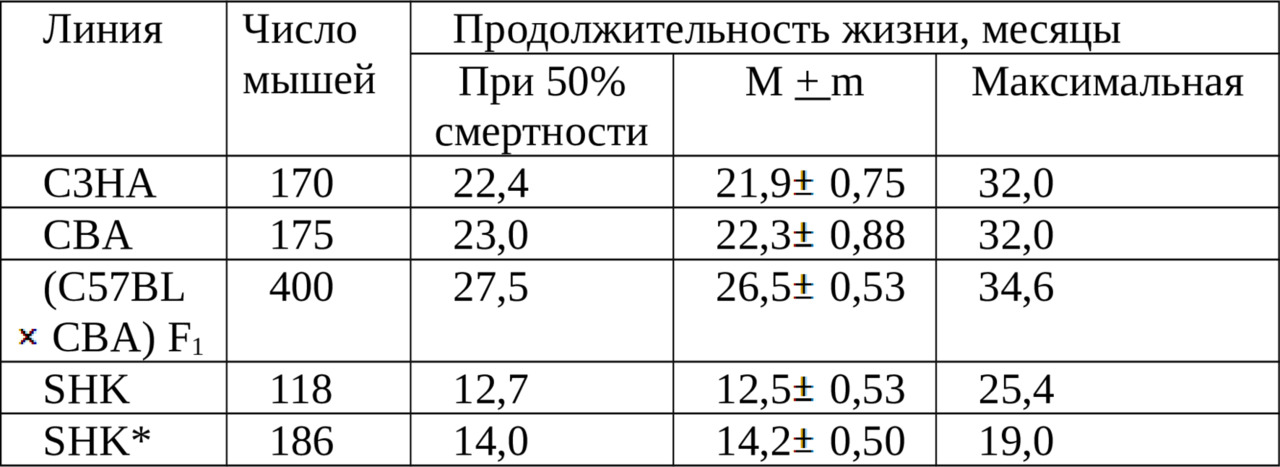
Таким образом, использование геропротектора (хлоргидрат 2-этил-6 метил-3-оксипиридин) приводит к замедлению процесса старения у мышей SHK в среднем в 1,8 раза (таблица 3 и рис. 3).

Экспериментальные данные о влиянии антиоксидантов на выживание экспериментальных животных нашли соответствующее отражение в литературе и, естественно, вызвали большое количество вопросов. Уместно остановиться на некоторых из них.
1-ый вопрос. Достоверен ли пролонгирующий эффект антиоксидантов?
Ответ. Известно, что при достаточной выборке (более 40 линейных мышей в группе) средняя продолжительность жизни (СПЖ) воспроизводится с точностью 3—4%, тогда как пролонгирующий эффект достигал 33%. Т.е. в этих случаях увеличение продолжительности жизни при действии антиоксиданта статистически достоверно и значимо. Для человека 30%-ое увеличение продолжительности жизни означало бы дополнительные 20 лет жизни.
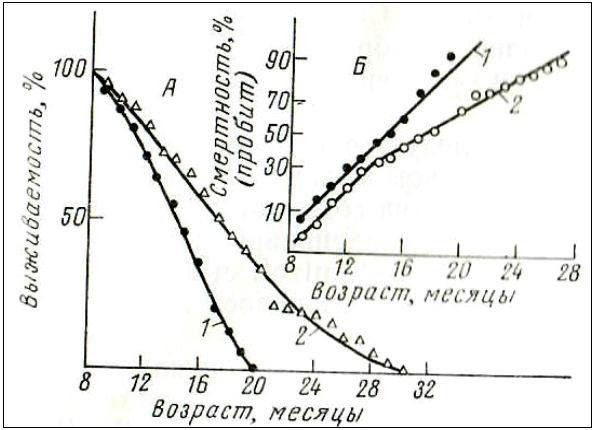
2-ой вопрос. Действует ли антиоксиданты только на короткоживущих (мутантных) мышей, предрасположенных к опухолям?
Ответ. Как видно, эмоксипин увеличивал среднюю (СПЖ) и максимальную продолжительность жизни (МПЖ) не только короткоживущих мышей (SHK), но и долгоживущих мышей (С3НА).
Т. е. СПЖ и МПЖ мышей SHK в контрольной группе были равны 15 и 19 месяцев, под воздействием антиоксиданта увеличились до 19 и 30 месяцев соответственно. Для мышей линии С3НА в контрольной группе СПЖ и МПЖ были равны 22 и 32 месяца, а под влиянием эмоксипина достигли 27 и 41 месяца. Тогда как, на высокораковую линию мышей AKR эта же доза эмоксипина на оказала никакого влияния.
3-й вопрос. Прямо или опосредованно действуют антиоксиданты?
«В эксперименте с ионолом параллельно увеличению средней продолжительности жизни снижалась масса тела мышей. Однако остается неясным, обусловлено ли специфическое действие антиоксидантов на длительность жизни непосредственно ослаблением свободнорадикальных реакций в тканях или оно опосредовано замедлением индивидуального развития на стадии роста у млекопитающих». Ответ. При анализе результатов экспериментов с ионолом видно, что различия веса подопытных и контрольных минимальны. Так по данным Хармана, средняя масса тела мышей контрольной группы составляла 31,6 г, а у подопытных, когда СПЖ увеличилась на 45%, масса тела была 29,2 г. Тогда как при сдерживающей рост диете масса тела контрольных животных была в 1,5 — 2 раза больше массы тела подопытных мышей.
К тому же, как потом оказалось, антиоксиданты сами по себе поливалентны и могут влиять одновременно на многие геронтологические процессы (более четырех механизмов или моделей старения).
На рисунке 4 показаны различные варианты воздействия антиоксидантов на кривые выживания:
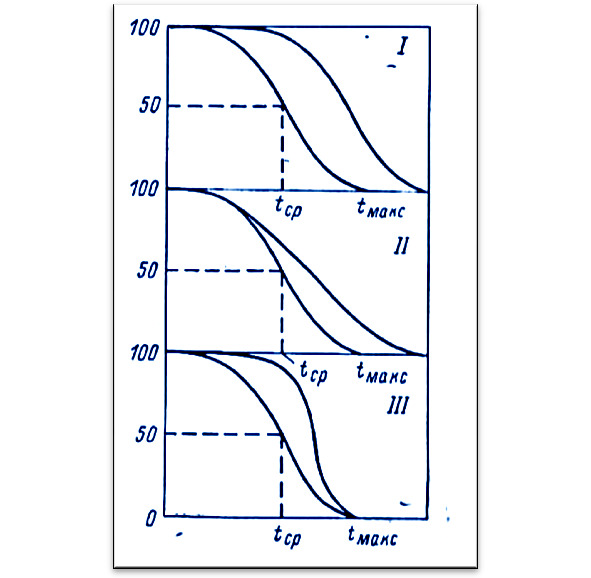
I — Равномерное замедление старения: равное увеличение средней и максимальной
продолжительности жизни (ПЖ).
II — Замедление старение с большой лаг-фазой: большее увеличение максимальной ПЖ.
III — Улучшение условий жизни: увеличение только средней ПЖ, а не максимальной ПЖ.
Вещества, которые замедляли старение (I и II вариант), стали называть геропротекторами.
Необходимым условием доказательства, что данное препарат действительно геропротектор, стало достоверное уменьшение смертности под его влиянием во всех возрастных группах экспериментальных животных. При этом условии не возникает вопрос о продлении жизни одних особей популяции за счёт укорочении жизни других. Особое значение приобрели работы, в которых установлен не только факт увеличения средней продолжительности жизни (СПЖ), но и максимальной продолжительности жизни (МПЖ). Изменение только СПЖ может быть связано не с процессом старения экспериментальных животных, а с изменением условий их жизни. Только после того, как установлен факт увеличения продолжительности жизни экспериментальных животных, серьезное значение приобретает изучение особенностей влияния этого вещества на отдельные системы организма. Легкость, с которой открывали все новые антиоксиданты-геропротекторы и свободно-радикальные патологии, создала бум вокруг антиоксидантов-геропротекторов.
Работы Хармана были процитированы более 5500 раз. В результате были поставлены проверочные, хорошо обставленные исследования на большой выборке (до 250 000 человек), которые показали, что антиоксидантные витамины в некоторых комбинациях не оказывают ожидаемого положительного влияния на продолжительность жизни человека, а в отдельных вариантах оказывают отрицательное действие. До сих пор эти работы широко цитируются в научной и популярной геронтологической литературе, хотя были опубликованы многочисленные разъяснения этих отрицательных эффектов антиоксидантных витаминов.
Уместно вспомнить, что научной теории, чтобы развиться, просто необходимы общественные проклятия, ненависть научной среды и осмеяние в прессе. Именно эти факторы обычно мобилизуют и вооружают серьезные идеи, провоцируют их на движение и победы [67]. Очевидно, что эти эксперименты ярко продемонстрировали, что иллюзия легкости обращения со свободно-радикальной тематикой возникла от недостаточности знаний в этой области, что свободно-радикальная теория еще нуждается в доработке и дальнейшем развитии. Т.е. возникла необходимость развития свободно-радикальной теории старения, которую мы решили назвать в новом варианте «антиоксидантная концепция старения». Что с самого начала во главу ставит не повреждение, а защиту от агрессоров. Следует подчеркнуть наиболее важные моменты:
— исправление ошибок интерпретации и логических несоответствий критических нападок противников антиоксидантов;
— связь свободно-радикальных механизмов старения с возрастным увеличением смертности от главных пато-геронтологических процессов (болезней системы кровообращения и злокачественных новообразований);
— стремление к пониманию старения именно человека. Так как механизмы старения человека часто отличаются от механизмов старения мышей и дрозофил.
С целью дальнейшего развития начатых работ в 1975 году на Секции химико-технологических и биологических наук АН СССР была сформирована комиссия по проблемам «Искусственного увеличения видовой продолжительности жизни людей». Председателем комиссии назначен академик Н. М. Эмануэль, его заместителем — Л. В. Комаров. Был составлен координационный план научных исследований по проблеме «Искусственное увеличение видовой продолжительности жизни людей» на 1976—1985 гг.
План охватывал свыше 50 тем, среди них такие, как «Эволюционные предпосылки возникновения генетической программы старения человека», «Роль наследственных факторов в собственно старческих и компенсаторно-старческих изменениях организма», «Биологические и социальные факторы долгожительства», «Изучение роли репартивных механизмов клетки» [71].
В 1975 году в Институте химической физики АН СССР была организована лаборатория «Количественной геронтологии» (первая геронтологическая лаборатория в АН СССР), которая развернула научную работу в сотрудничестве с Центральной клинической больницей УД АН СССР (заведующий Т. Л. Наджарян, совмещающий заведование отделением реанимации ЦКБ УД СССР, и его заместитель В. Б. Мамаев).
С самого начала в основу наших исследований был положен количественный подход в отличии от Киевского института геронтологии МЗ СССР. Мы основной акцент в нашей работе делали на математическое описание реальных процессов, которые можно объективно установить без теоретических предположений и допущений. Кроме того под руководством академика Н. М. Эмануэля и его заместителем Т. Л. Наджаряна начал свою работу московский научный семинар «Фундаментальные проблемы биологии старения», материалы которого были опубликованы в трех сборниках «Итоги науки и техники ВИНИТИ» [51, 7, 8].
«Кому нужны старые мыши?»
С этим вопросом или утверждением Н.М. открывал совместную работу лаборатории количественной геронтологии с Центральной клинической больницей УД АН СССР.
Первый подход к замедлению старение человека исследовать репликативное старение клеток человека в культуре.
И следующим этапом развития было изучение влияния антиоксидантов-геропротекторов на клетки человека, что возможно только в условиях in vitro. К этому времени были три группы работ, которые указывали на возможность такого влияния.
Во-первых, было показано [76, 89] влияние парциального давления кислорода на рост, метаболизм и клеточный цикл диплоидных клеток человека в культуре. В клеточном цикле существуют периоды, чувствительные к повышению концентрации кислорода и супероксидных радикалов. При повышении концентрации кислорода деление нормальных клеток подавлялось [88, 96].
Во-вторых, при нормальном парциальном давлении кислорода введение в культуральную среду антиоксидантов, наоборот, стимулировало деление клеток [2].
В-третьих, при сопоставлении продолжительности жизни клеток в кровяном русле c активностью СОД [57, 58], уровнем генерации супер-оксидных радикалов в гранулоцитах [75], тромбоцитах [94], эритроцитах [95] и лимфоцитах [90] видно, что гранулоциты обладают наименьшей активностью СОД, экзогенно генерируют супер-оксидный радикал и живут только 12—14 часов. А лимфоциты при высокой активности СОД экзогенно не генерируют супер-оксидные радикалы и живут 400 и более дней. Т.е. продолжительности жизни клеток крови во многом определяется степенью окислительного «стресса».
Влияние антиоксидантов-геропротекторов на репликативное старение диплоидных клеток человека [56].
Монослойное культивирование сопровождается закономерным изменением числа делящихся клеток в течение каждого пересева: в логарифмической фазе роста культуры, с 1-го по 3-ий день роста, — максимальное число делящихся клеток.
Монослой клеток образуется к 3-4-му дню культивирования. К 9-му дню культуры переходят в стационарную фазу, когда число делящихся клеток незначительно и плотный пласт фибробласто-подобных параллельно-ориентированных клеток.
В некоторых культурах в эти сроки клетки располагаются в несколько слоев — один слой перпендикулярно к другому. Вероятно, различные фазы роста после пересева моделируют состояние клеток в различных популяциях организма: возобновляющихся, растущих или стабильных популяциях.
Добавляя антиоксиданты на различных фазах роста культуры, мы воздействовали на клетки в состоянии деления и покоя. Культура в логарифмической фазе роста подобна быстро обновляющимся клеточным популяциям, а культура в стационарной фазе — медленно обновляющимся популяциям клеток взрослого организма.
При воздействии антиоксидантов на культуры лаг-фазы и фазы логарифмического роста, т.е. при добавлении эмоксипина в момент пересева число меченых 3Н-тимидином клеток достоверно увеличивается. Для изучения влияния эмоксипина на пролиферацию клеток только в экспоненциальной фазе роста культуры препарат вводили в среду через 24 часа после пересева и фиксировали еще через 24 часа, т.е. через 48 часов после пересева. В этом случае препарат даже в высокой концентрации не влиял на пролиферацию. Число меченых клеток составило в контрольных культурах Si±σ = 92,7±1,4% (11 пассаж).
В отличие от этого, введение эмоксипина в культуры, находящиеся в стационарной фазе, существенно стимулировало пролиферацию клеток. Культура стационарной фазы характеризовалась почти полным прекращением пролиферации клеток [77].
Добавление эмоксипина приводило к существенному увеличению числа меченых клеток (таблица 4).
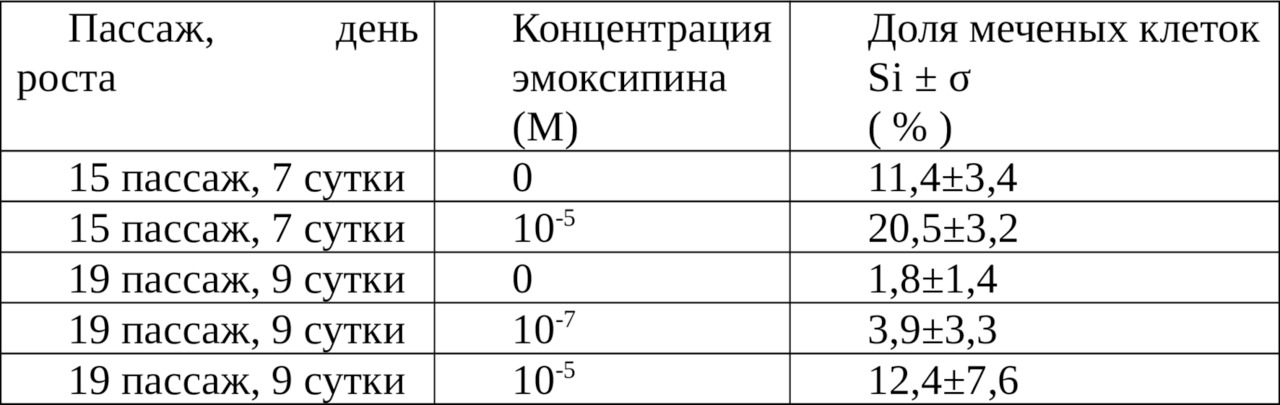
Относительно контрольных культур, которые характеризуются малой величиной числа меченых клеток (в соответствии с глубиной выраженности стационарной фазы — 11,4±3,4% и 1,8±1,4%), это увеличение выглядит более значительным (до 20,5±3,2 и 12,4±7,6% соответственно), чем в пролиферирующих культурах. Таким образом, относительная величина стимуляции возрастает с уменьшением числа ДНК-синтезирующих клеток в контрольной культуре. Под влиянием антиоксиданта число меченых клеток в стационарной фазе увеличивается примерно на 10%.
Известно, что в процессе пассирования культур диплоидных клеток человека происходит характерное для старения нормальных клеток снижение числа меченых 3Н-тимидином клеток [82, 86]. Так, в наших исследованиях на 13-м и 45-м пассажах через 48 часов культивирования число меченых клеток было 80,8±5,5% и 29,0±2,2%.
Мы изучали также влияние эмоксипина на пролиферацию ДКЧ при постоянном присутствии препарата (10—5 М эмоксипина, на протяжениии 25 суток)
на протяжении семи пассажей (с 36-го до 42-й пассаж). В этих условиях эмоксипин замедлял снижение числа клеток, синтезирующих ДНК, по мере культивирования, то есть тормозил характерное для пролиферативного старения диплоидных клеток человека снижение пролиферации. Так, в контрольных культурах с 36-го по 42-ой пассаж доля клеток, синтезирующих ДНК, снизилась с 88,7±3,7% до 66,0±8,0%. Тогда как в присутствии 10—5 М эмоксипина доля клеток, синтезирующих ДНК, на 42-ом пассаже составила только 83,6±1,7%.
Следует подчеркнуть, что в данном эксперименте при постоянном присутствии антиоксиданта в течение 7 пассажей не наблюдали изменений кариотипа и морфологии клеток. Что позволило сделать важный вывод об отсутствии токсического действия эмоксипина в дозах 10—5М на нормальные клетки человека.
Эти данные хорошо согласуются с фактами стимуляции антиоксидантами митотической активности клеток паренхимы печени и клеток эпителия тонкой кишки мышей in vivo [37, 59, 60]. Добавление в пищу мышам эмоксипина стимулировало кроветворение после кровопотери у мышей старших возрастных групп [69, 70]. Показано, что при введении антиоксидантов в дозах, увеличивающих антиокислительную активность (АОА), происходит ускорение деления клеток и уменьшения среднего времени генерации.
Напротив, антиоксиданты в дозах, уменьшающих антиокислительную активность, тормозят размножение клеток, тормозя их вступление в митоз [3, 4]
Для объяснения этих экспериментальных данных было выдвинуто несколько гипотез [5, 83, 97, 98]. Гипотезу Е. Б. Бурлаковой [5, 6, 79] можно проиллюстрировать схемой, составленной по её работам и представленной на рисунке 5.
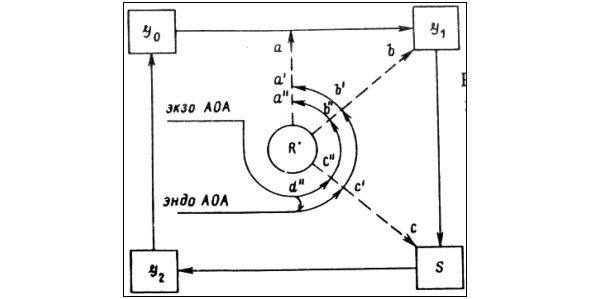
(G0, G1, S, G2). экзо АОА — экзогенные антиоксиданты, эндо АОА — эндогенные антиоксиданты, R• — свободные радикалы.
Действие свободных радикалов (R•) в наиболее чувствительные фазы клеточного цикла может подавить или полностью блокировать деление клеток (a, b, c). Эндогенные антиоксиданты, характеризующиеся высокой антиокислительной активностью, ингибируют свободно-радикальные процессы и снимают этот блок (a’, b’, c’). Той же способностью обладают и экзогенные антиоксиданты (a», b», c»). Кроме того введение экзогенных антиоксидантов способствует увеличению окисляемости эндогенных липидов и уменьшению эндогенной антиокислительной активности (d»). Таким образом, биологическая система поддерживает оптимальный уровень антиокислительной активности. Материальным субстратом, в котором осуществляется этот процесс, являются липидные образования (мембраны и липопротеиды).
Таким образом, на основании полученных результатов и литературных данных можно предположить, что одним из механизмов замедления старения животных антиоксидантами является стимуляция к делению части клеточной популяции, особенно тех, которые находились в состоянии покоя (G0-фазе). Возможно антиоксиданты-геропротекторы в оптимальной концентрации восстанавливают состояние наружной мембраны, необходимое для реализации действия факторов роста, запускающих деление клетки.
В 2009 году наступил новый период развития геронтологии после вручения Нобелевской премии за открытие механизмов защиты хромосом с помощью теломер и теломеразы, существование которых еще в 70-ые годы теоретически предсказал А. М. Оловников. Казалось, что открыт новый и единственный механизм старения в противовес свободно-радикальной теории старения. По мере исследований выяснилось, что эти механизмы существуют одновременно и как бы дополняют друг друга.
Что же дальше?
Несмотря на очевидные достижения биологии исчерпывающего ответа на вопрос: «Чем отличаются зародышевые клетки от соматических клеток?» пока нет. Ответ на этот исходный вопрос, давно поставленный Вейсманом, во многом определит дальнейшее развитие геронтологии.
Большие надежды возложены на развитие методической базы регенеративной медицины.
Список приведенной литературы и важных публикаций по теме.
1. Бурлакова Е. Б., Гаинцева В. Д., Слепухина Л. В., Храпова Н. Г., Эмануэль Н. М. Антирадикальная активность и радиозащитные свойства ингибиторов радикальных реакций. // Доклады АН СССР, 1964. Т. 155, №6. С. 1398—1400
2. Бурлакова Е. Б., Дубинская Н. И., Коперина Е. В., Пальмина Н. П. // Биофизика, 1966. Т.11, №6, С.1008—1012.
3. Бурлакова Е. Б., Пальмина Н. П. // Биофизика, 1966. Т.11. №2. С.258—262.
4. Бурлакова Е. Б., Пальмина Н. П. // Биофизика, 1967. Т.12. №6. С.1032—1036.
5. Бурлакова Е. Б. // Биофизика, 1967. Т.12. №1. С.82—88.
6. Бурлакова Е. Б. // В сб.: «Биоантиоксиданты в лучевом поражении злокачественном росте». М: Наука, 1975. С.172—199.
7. Бурлакова Е. Б. и Наджарян Т. Л. (ред.) Итоги науки и техники. Серия «Общие проблемы биологии». Том 5. Биологические проблемы старения. Замедление старения антиоксидантами. М.: ВИНИТИ, 1986.
8. Бурлакова Е. Б. и Гаврилов Л. А. (ред.) Итоги науки и техники. Серия «Общие проблемы биологии». Том 6. Популяционная геронтология. М.: ВИНИТИ, 1987.
9. Эмануэль Н. М., Семенов Н. Н. Кинетический метод исследования промежуточных продуктов автокаталитических реакций. // Доклады АН СССР, 1940. Т.28. №3. С. 220—223.
10. Эмануэль Н. М., Павлов Д. С., Семенов Н. Н. SO — промежуточный продукт реакции окисления сероводорода. // Доклады АН СССР, 1940. Т.28. №7. С. 619—621.
11. Эмануэль Н. М. Макроскопические стадии, особая роль начального периода и механизм действия ингибиторов и положительных катализаторов в цепных реакциях. // Доклады АН СССР, 1954. Т.95. №3. С. 603—606.
12. Эмануэль Н. М. Стимулирование цепных разветвленных реакций воздействием проникающих излучений в начальный период развития процесса // Докл. АН СССР, 1956. Т. 111. №6. С. 1286—1289.
13. Эмануэль Н. М. Цепная теория действия ингибиторов в процессах окисления пищевых жиров. // В кн.: Тезисы докладов на Всесоюзном научно-техническом совещании по вопросам окислительных процессов в пищевых жирах и методов защиты жиров от окисления и прогоркания. М., 1957, с. 3.
14. Эмануэль Н. М., Бурлакова Е. Б., Дзантиев Б. Г., Сергеев Г. Б. К вопросу о значении цепного окисления жиров под действием излучения для развития лучевого поражения. // В кн.: Биохимические и физико-химические основы биологического действия радиации (25—28 февраля 1957 г.): Тезисы докладов. М., 1957, с. 10—12.
15. Эмануэль Н. М., Липчина Л. П. Лейкоз у мышей и особенности его развития при воздействии ингибиторов цепных окислительных процессов. // Доклады АН СССР. 1958. Т. 121. Вып. 1. С. 141—144.
16. Эмануэль Н. М. Роль свободных радикалов в радиобиологических процессах и некоторые новые возможности разработки средств против лучевого поражения. // В кн.: Первичные механизмы биологического действия ионизирующих излучений. (Труды МОИП. Т. 7.) М.: 1963. С 73—83.
17. Бурлакова Е. Б., Гаинцева В. Д., Слепухина Л. В., Храпова Н. Г., Эмануэль Н. М. Антирадикальная активность и радиозащитные свойства ингибиторов радикальных реакций. // Доклады АН СССР, 1964. Т. 155, №6. С. 1398—1400
18. Эмануэль Н. М. Можно ли жить дольше? // Лит газета. 1968. 28 августа. С. 13.
19. Эмануэль Н. М. Физико-химические аспекты процесса старения и проблема увеличения продолжительности жизни. // Труды IX международного конгресса геронтологов. Киев, 2—7 июля 1972. Рефераты докладов и выступлений. Киев: 1972. Т. 1. С.295.
20. Emanuel N.M. Die Inhibitoren des Alterns. // Ideen des exakten Wissen, 1973. N 7. S. 456—462. (Эмануэль Н. М. Ингибиторы старения. // Идеи точного Знания, 1973. N 7. С. 456—462).
21. Эмануэль Н. М. В поисках преград старению. // Ленинское знамя, 1973, 8 июля. // Заря Востока, 1973, 8 июля.
22. Эмануэль Н. М. Можно решать любую задачу. // Литературная газета, 1973. 29 августа. С 11.
23. Эмануэль Н. М. Некоторые молекулярные механизмы и перспективы профилактики старения // Известия АН СССР. Серия биологическая, 1975. №4. С.785—794.
24. Эмануэль Н. М. Свободные радикалы и старение. // Геронтология и гериатрия 1975: Ежегодник «Биологические возможности увеличения продолжительности жизни». Киев: 1976. С. 103—110.
25. Emanuel N.M. Free radicals and the action of inhibitors of radical processes under pathological states and ageing in living organisms and in man. // Quarterly Reviews of Biophysics. Amsterdam: 1976. V. 9. N 2. P. 283—308.
26. Эмануэль Н. М., Обухова Л. К., Бунто Т. В., Дьякова В. В. Кинетические закономерности выживаемости и экспериментальное определение скорости старения. // Известия АН СССР. Серия биологическая, 1976. №6. С.789—794.
27.Эмануэль Н. М., Обухова Л. К., Смирнов Л. Д., Бунто Т. В. Эффект увеличения продолжительности жизни в эксперименте при воздействии хлоргидрата 2-этил-6-метил-3-оксипиридина. // Доклады АН СССР, 1976. Т. 226. №4. С. 961—963.
Бесплатный фрагмент закончился.
Купите книгу, чтобы продолжить чтение.