
Бесплатный фрагмент - В поисках прародины индоевропейцев
Теория сдвигов согласных звуков
Глава I. Использование данных ДНК для реконструкции истории человечества
1.1. Новые возможности для изучения древней истории человечества
Достижения в области генетических исследований, произошедшие за последнее десятилетие, дают возможность восстановить историю человечества на основании информации, зашифрованной в наших генах. Новые технологии в области анализа ДНК человека позволяют не только установить родство между определенными популяциями современных людей, но и проводить исследования ДНК древних захоронений, найденных при раскопках поселений первобытных людей. Изучение информации, содержащейся в ДНК современных людей, позволяет заглянуть вглубь тысячелетий и проследить миграции наших далеких предков в ходе их расселения по территории нашей планеты.
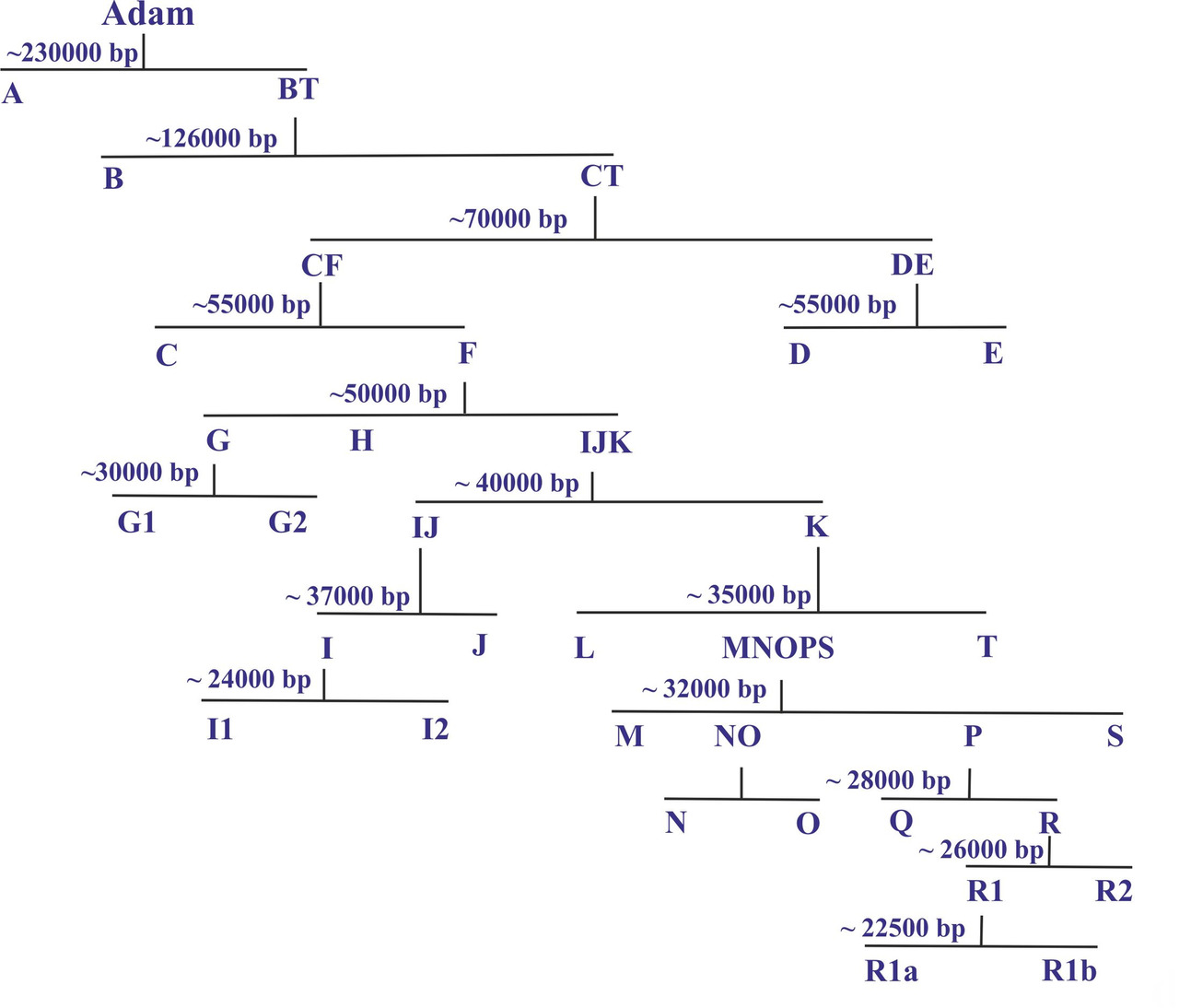
Реконструкция последовательности разделение древних родов по мужской линии, начиная от гипотетического Адама, является одним из наиболее важных результатов исследований ДНК человека. На основании этих данных было построено дерево y-гаплогрупп, которое в упрощенном виде может быть представлено следующим образом [1] [2] (рис.1).
Возможность определения маршрутов миграций древних людей позволяет выработать новый подход к решению проблем, связанных с локализацией территорий возникновения протоязыков и созданием моделей их распространения на определенной территории. Одной из наиболее интересных проблем современной лингвистики является создание модели распространения индоевропейских языков и установление прародины индоевропейцев. До сих пор все известные научные модели строились на попытках установления связей между распространением индоевропейских языков и распространением определенных археологических культур или технологических инноваций (изобретение колеса и первых транспортных средств).
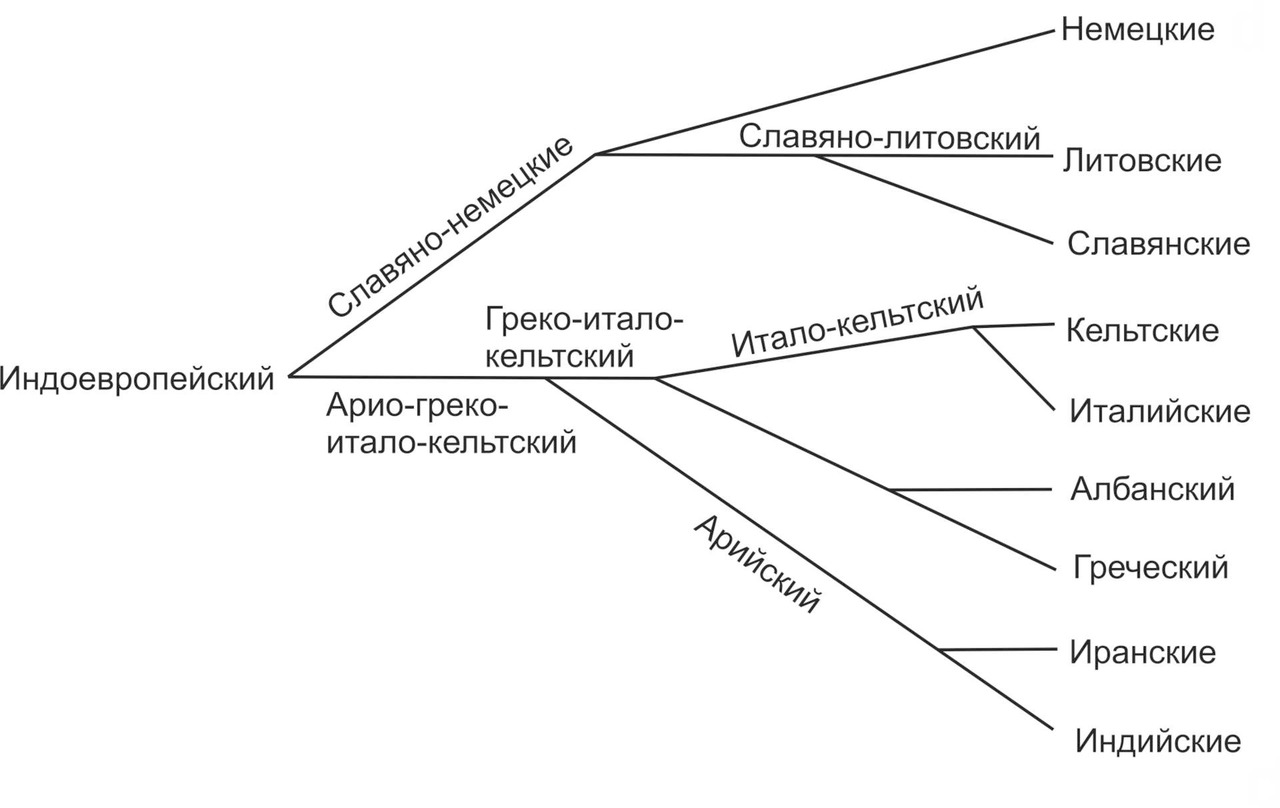
Основоположником сравнительно-исторического языкознания считается известный британский филолог и переводчик Уильям Джонс. Джонс был первым, кто обратил внимание на сходство между европейскими языками и санскритом. Свои наблюдения Джонс изложил в книге «Санскритский язык» (The Sanscrit language), которая увидела свет в 1786 году. С тех пор прошло уже больше двухсот лет. За это время индоевропеистика проделала большой путь, однако главная загадка, касающаяся установления прародины носителей индоевропейских языков, все еще считается неразгаданной. Кроме того, до сих пор остается открытым вопрос, касающийся филогении индоевропейских языков. Одной из наиболее известных схем разделения основных ветвей индоевропейских языков является схема немецкого лингвиста Августа Шле́йхера (рис.2).
Большинство современных лингвистов связывает распространение индоевропейских языков на территории Евразии с расселением носителей y-гаплогруппы R1a. На сегодняшний день накоплены значительные данные по «европейским» и «азиатским» субкладам y-гаплогруппы R1a. Эти данные указывают на существование корреляции между распространением субкладов R1a и распространением различных ветвей индоевропейских языков на территории Евразии. Сторонники теории распространения индоевропейских языков носителями y-гаплогруппы R1a считают, что филогения субкладов y-гаплогруппы R1a и высокий процент мужчин-носителей данной y-гаплогруппы среди популяций, говорящих на индоевропейских языках, делают носителей данной y-гаплогруппы наиболее вероятными распространителями индоевропейских языков на территории Евразии.
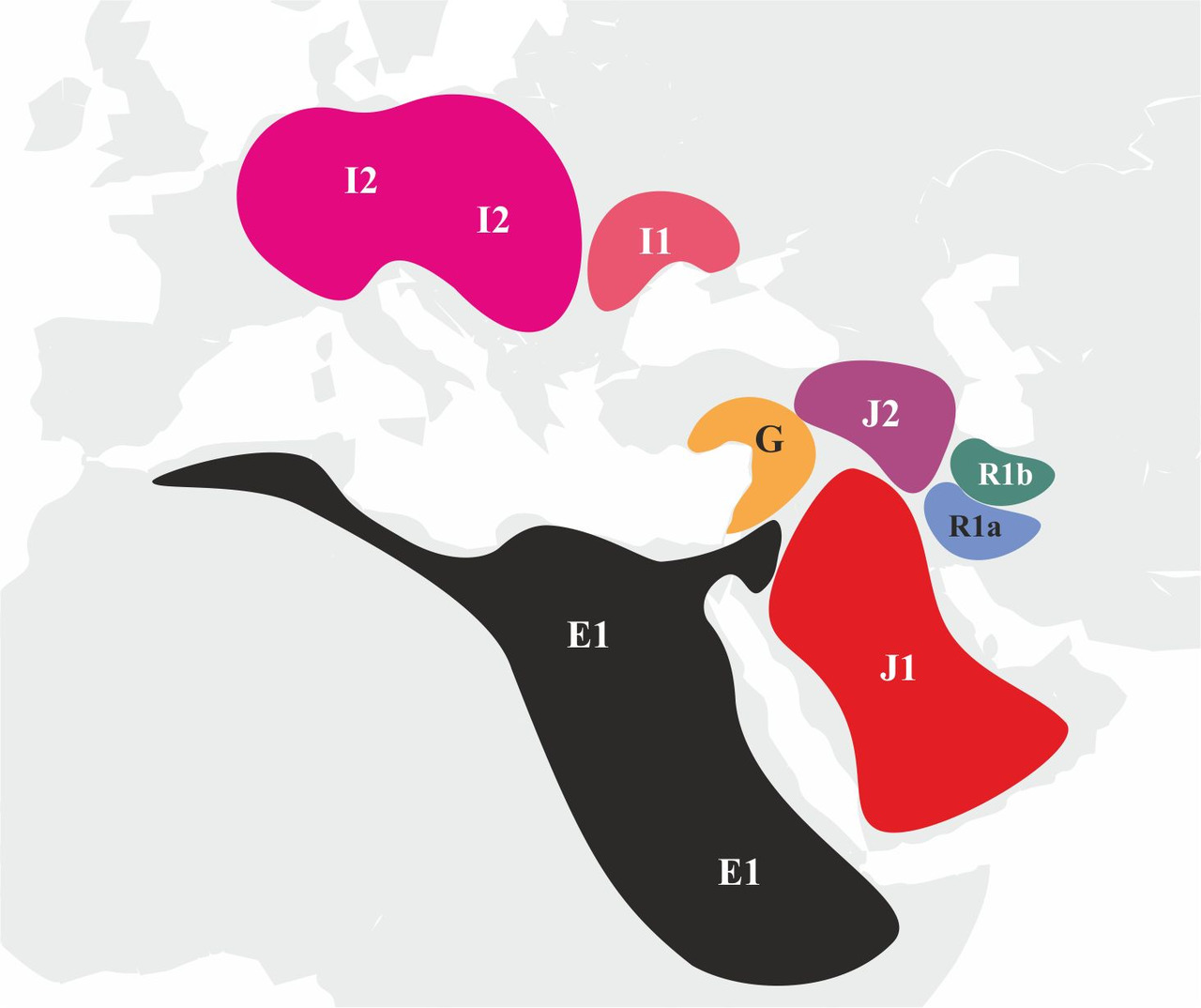
В некоторых случаях попытки ассоциировать распространение языков, принадлежащих к какой-либо языковой семье, с миграцией носителей определенной y-гаплогруппы или даже субклада являются вполне оправданными. Например, подавляющее большинство коренных жителей Северной и Южной Америки являются носителями субклада Q1a3, что дает основание считать, что распространение языков индейцев Северной и Южной Америки связано с носителями именно этого субклада. Схожая ситуация наблюдается с сино-тибетскими языками: у популяций, говорящих на сино-тибетских языках, чаще всего встречается y-гаплогруппа O. С народами, живущими на территории Европы, все гораздо сложнее. Современные популяции европейцев являются результатом смешения носителей различных y-гаплогрупп (рис.3).
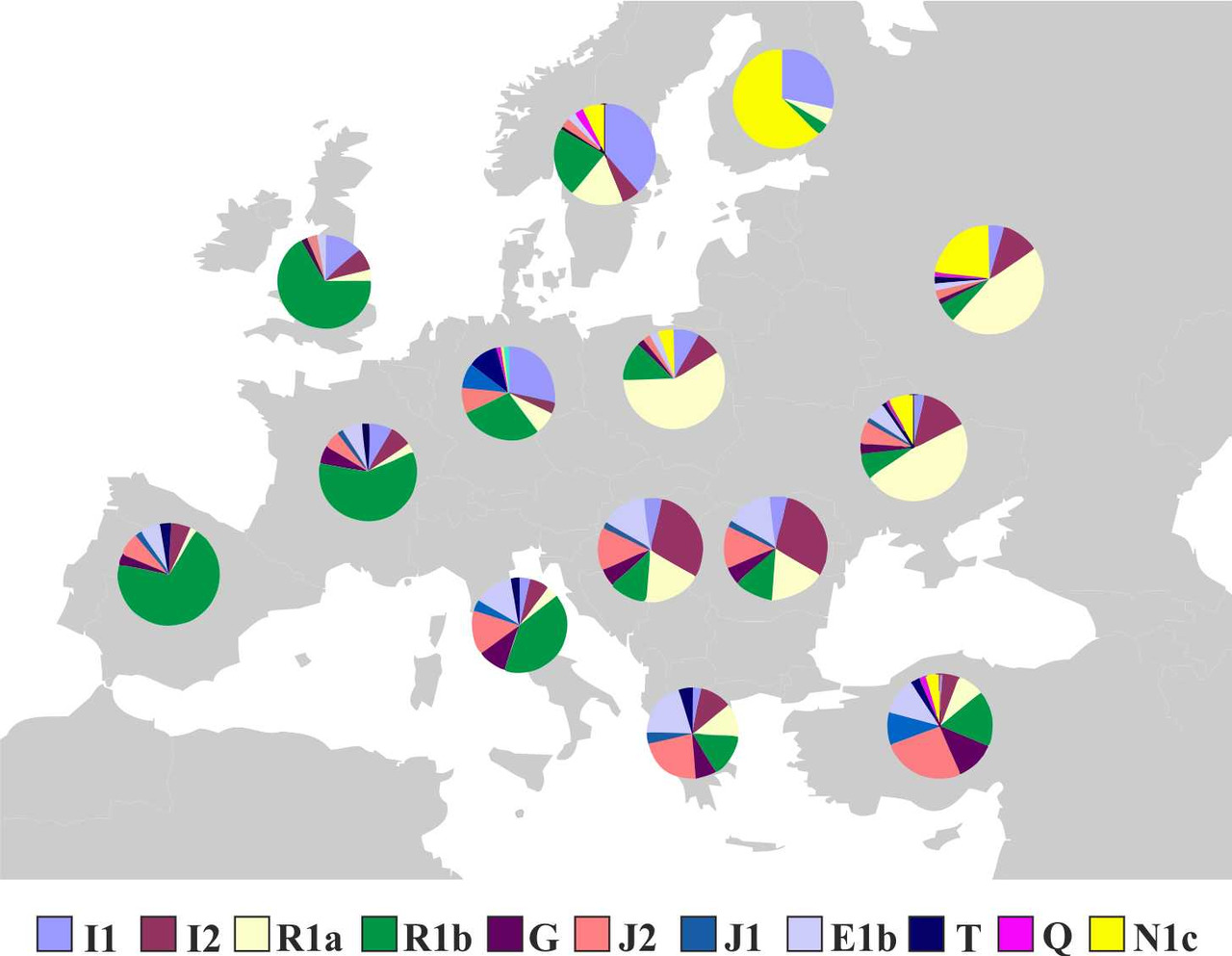
Хотя среди них есть доминирующие y-гаплогруппы, например, в западной Европе доминируют носители y-гаплогруппы R1b, а в Центральной и Восточной Европе носители y-гаплогруппы R1a, строгого соответствия между распределением y-гаплогрупп и распределением языков, принадлежащих к определенным языковым семьям, все же не наблюдается. Например, у басков процент носителей y-гаплогруппы R1b составляет около 95%, но при этом, в отличие от своих индоевропейских соседей, они говорят на баскском языке, который не относится к индоевропейской языковой семье. В структуре y-гаплогрупп населения Венгрии совокупная доля носителей y-гаплогрупп R1a и R1b составляет около 50%, но при этом венгерский язык относится к уральской языковой семье, а не индоевропейской. Похожая ситуация наблюдается и в Азии: киргизы являются тюрками, хотя большая часть мужского населения (~75%) является носителями у-гаплогруппы R1a.
Принимая во внимание тот факт, что все современные европейские популяции появились в результате смешения представителей различных y-гаплогрупп, будет обоснованным предположить, что и их языки по своей сути тоже являются гибридами. Следовательно, возникает объективная необходимость в создании методики, которая бы позволяла провести анализ языка интересующей нас популяции для оценки вероятного вклада носителей определенных y-гаплогрупп при его генезисе. Другими словами, существует объективная необходимость в создании «моста», соединяющего ДНК генеалогию и лингвистику.
1.2. Основы ДНК генеалогии
ДНК генеалогия основана на возможности установления очередности и времени появления уникальных молекулярно-генетических меток. Такие метки периодически возникают в y-хромосомах и передаются от отца к сыну. В качестве молекулярно-генетических меток в ДНК генеалогии используются мутации (однонуклеотидный полиморфизм), которые по-английски называются single nucleotide polymorphism (далее — снип). Последовательность возникновения снипов в разных y-гаплогруппах была установлена путем сравнения между собой результатов анализов, полученных при тестировании мужчин, проживающих в разных регионах нашей планете. На основе этих данных ученым удалось построить филогеническое дерево y-гаплогрупп.
Периодичность возникновения снипов в y-хромосомах была рассчитана на основании данных, полученных при научных исследованиях полных сиквенсов y-хромосом. Оказалось, что в среднем одна мутация (снип) возникает в у-хромосоме один раз в три поколения (одно поколение — 25—30 лет), т.е. одна мутация происходит в среднем через каждые 75—90 лет. Данное свойство дает возможность использовать мутации для расчета возраста y-гаплогрупп или отдельных субкладов внутри y-гаплогруппы. Например, снип R1a-M417, который присутствует в у-хромосоме примерно 98% всех ныне живущих мужчин, имеющих y-гаплогруппу R1a, возник примерно 8500 лет назад. Это означает, что общий предок мужчин, являющихся носителями снипа R1a-M417, жил примерно 8500 лет назад. Чтобы понять, где жил этот предок, необходимо проанализировать данные географического распределения снипов, находящихся ниже и выше этого снипа. В связи с тем, что многие снипы y-гаплогруппы R1a были найдены сравнительно недавно и в научных исследованиях не тестировались, информация о распределении различных субкладов R1a на территории Евразии далека от того, чтобы считаться полной. Соответственно, при определении места возникновения снипа R1a-M417 мы должны ориентироваться не на снипы, которые расположены непосредственно ниже и выше снипа R1a-M417, а на снипы, по которым есть данные, полученные при проведении научных исследований. Носители реликтовых субкладов R1a (субклады, расположенные выше снипа R1a-M417) встречаются достаточно редко. Согласно данным научных исследований [3] из 24 выявленных носителей R1a-M420 (xSRY10831.2) 18 были найдены в Иране и 3 в Восточной Турции, а из шести найденных R1a1-SRY10831.2* (xM417) 5 человек были из Ирана и один кабардинец из России. В тоже время носители субклада R1a1-M417*, у которых отсутствуют известные снипы ниже снипа R1a-M417, были найдены только на территории Европы. Таким образом, на основании очередности возникновения снипов можно предположить, что снип R1a1-M417, скорее всего, возник где-то на границе между Ираном и Южным Кавказом, а до этого носители y-гаплогруппы R1a жили где-то на территории Иранского нагорья.
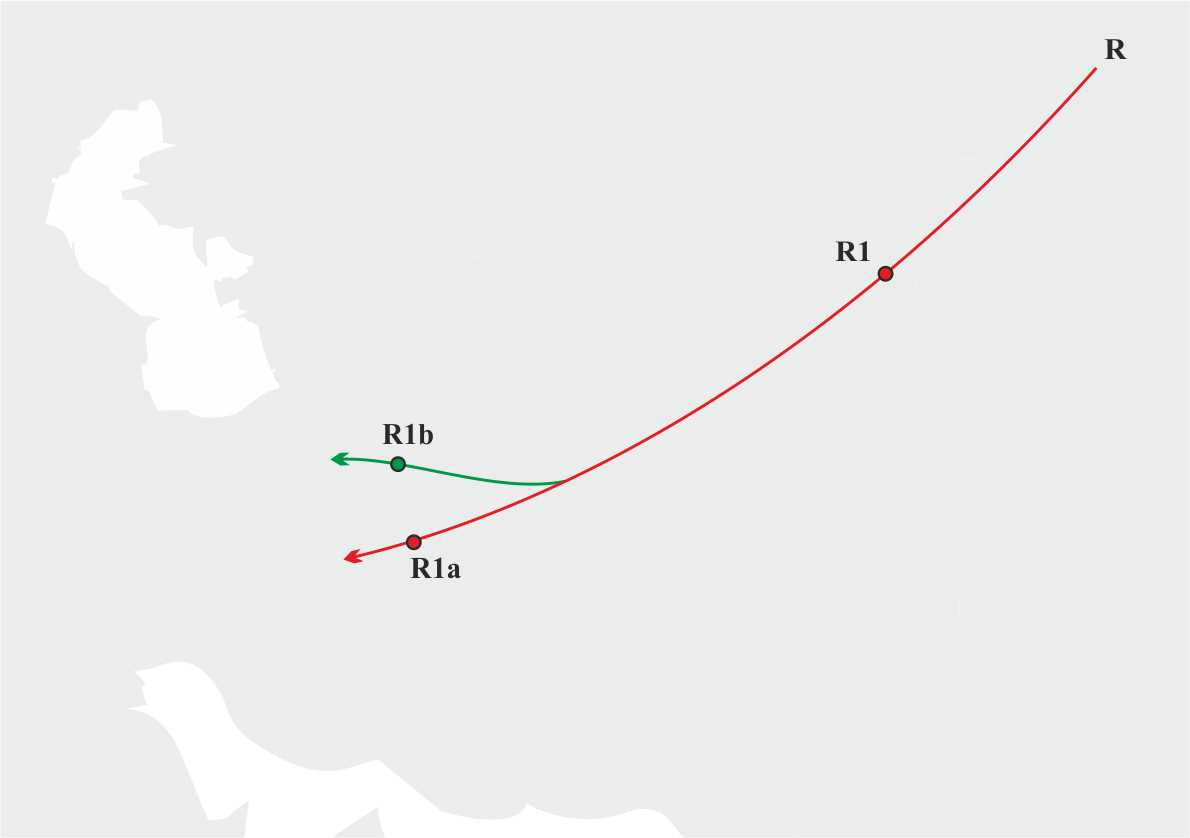
Таким же образом можно восстановить маршруты миграции наших более отдаленных предков. Например, предки большей части мужского населения сегодняшней Европы (носители y-гаплогрупп R1a и R1b) пришли на территорию Европы из Азии. Y-гаплогруппа R возникла где-то на территории Сибири примерно 30 тысяч лет назад. Исследования человеческих останков, найденных на позднепалеолитической стоянке в Сибири у нынешнего села Мальта Усольского района Иркутской области [4], показали, что останки принадлежали носителю y-гаплогруппы R. Y-гаплогруппа R1, вероятно, возникла так же на территории Сибири примерно 27 500 лет назад. С началом ледникового периода одна из групп носителей y-гаплогруппы R1 мигрировал из Сибири в Среднюю Азию, где примерно 22 000 лет назад произошло разделение на y-гаплогруппы R1a и R1b. Далее носители y-гаплогрупп R1a и R1b продолжили миграцию в юго-западном направлении на территорию Иранского нагорья (рис.4).
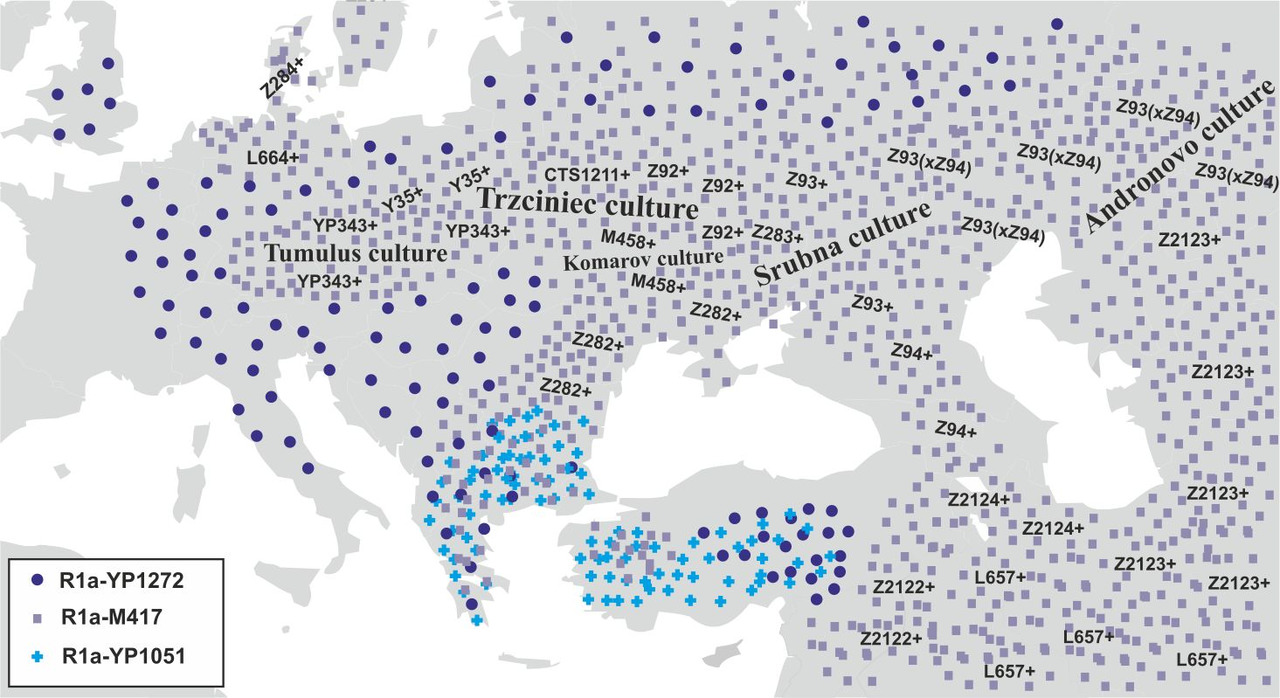
Для определения основных маршрутов миграций носителей y-гаплогруппы R1a с территории Иранского нагорья в Европу необходимо проанализировать распределение основных субкладов данной y-гаплогруппы. Таких субкладов всего три: R1a-YP1272 — возник примерно 14300 лет назад, R1a-YP1051- возник примерно 8500 лет назад и R1a-M417 — возник примерно 8500 лет назад. Несмотря на то, что носителей субкладов R1a-YP1272 и R1a-YP1051 [2] (рис.5) найдено не так уж и много, тем не менее картина распределения выявленных при тестировании носителей данных субкладов дает возможность определить маршруты миграций всех трех основных субкладов y-гаплогруппы R1a.
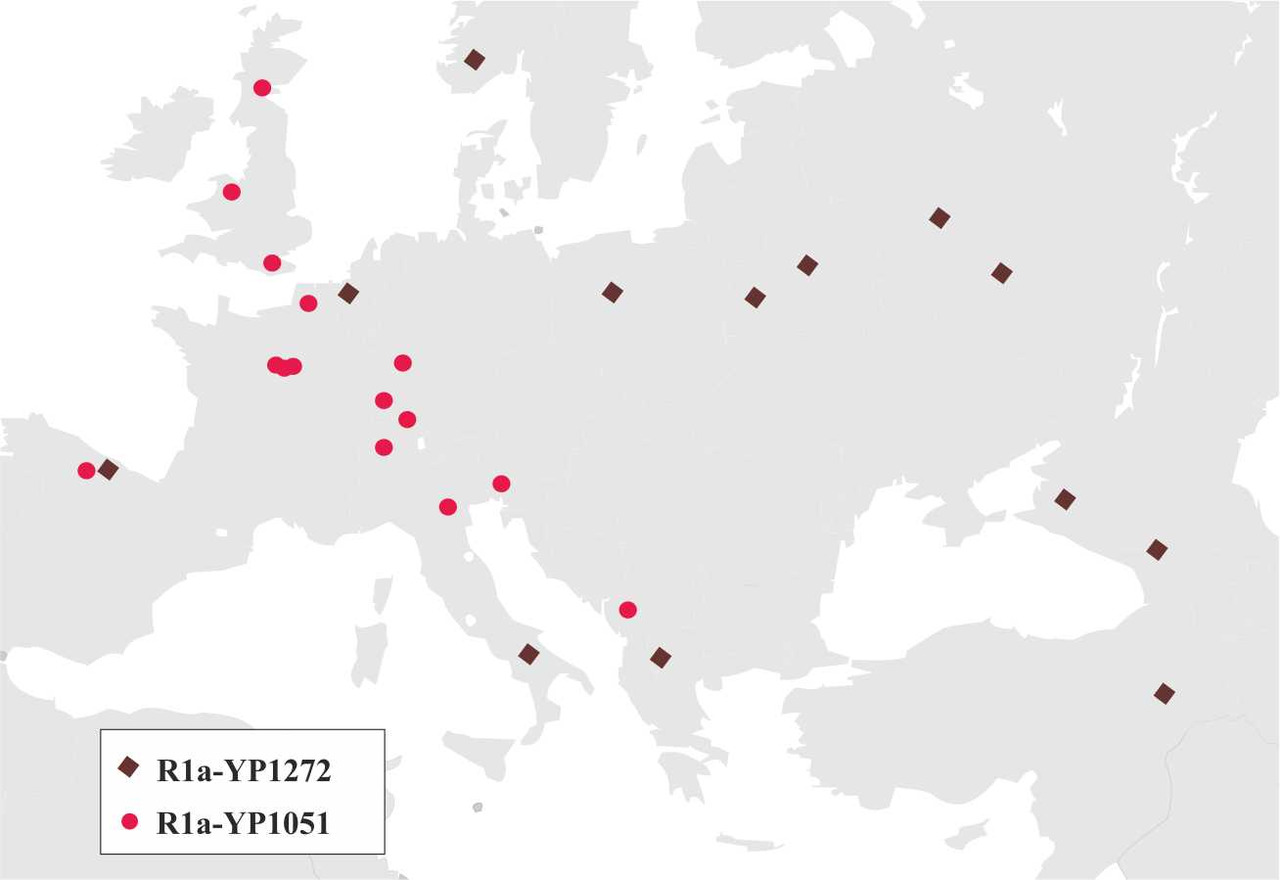
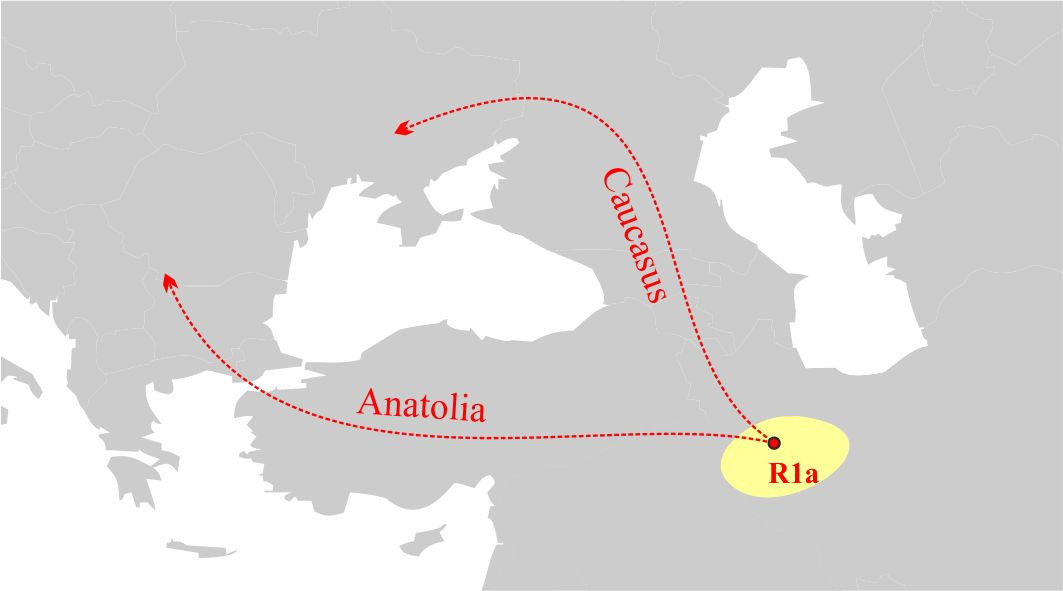
Анализ распределения носителей субкладов R1a-YP1272 и R1a-YP1051 на территории Европы дает основания предположить, что миграция носителей y-гаплогруппы R1a на территорию Европы проходила двумя основными маршрутами: первый маршрут — через территорию Анатолии на Балканский полуостров, а второй — через Кавказские горы между Черным и Каспийским морями (рис.6).
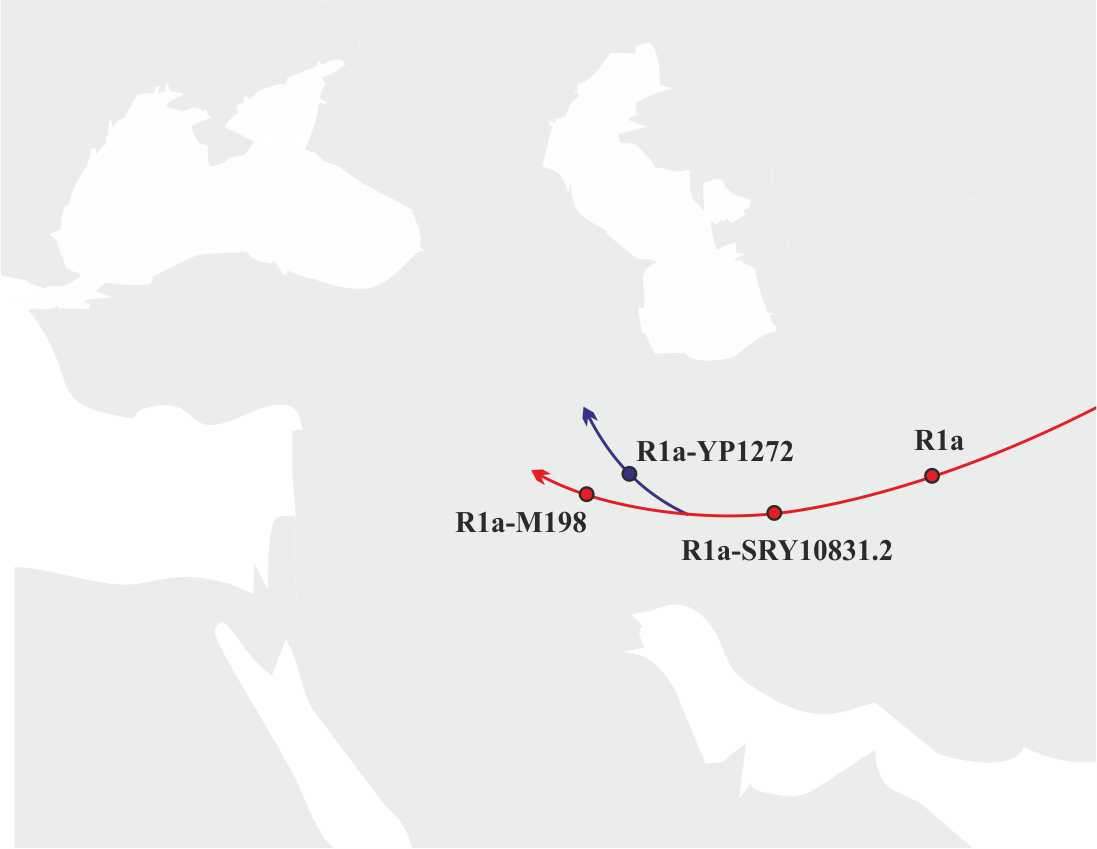
Учитывая данные о распределении носителей субкладов, лежащих ниже снипа R1a-M417 (снипы R1a-Z283 (xZ282) и R1a-Z93 (xZ94)), носители субклада R1a-M417, очевидно, мигрировали на территорию Европы по тому же маршруту, что и носители субклада R1a-YP1272, т.е. через Кавказские горы. На основании этой информации можно достаточно точно реконструировать основные маршруты и этапы миграции носителей y-гаплогруппы R1a на территорию Европы:
а) Примерно 14300 лет назад происходит разделение y-гаплогруппы R1a на субклады R1a-YP1271 и R1a-M198 (Иранское нагорье) (рис.7).
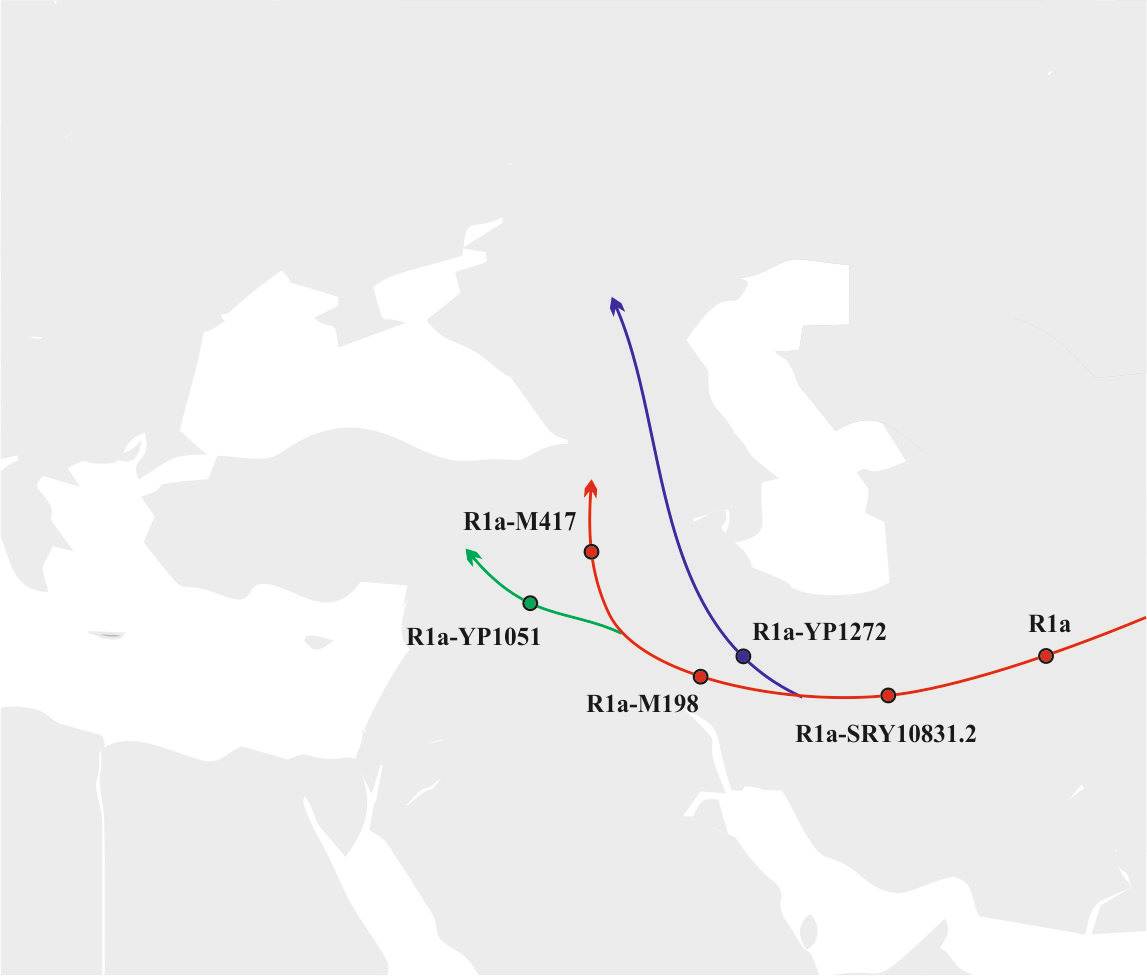
б) Примерно 8500 лет назад где-то на территории Месопотамии носители субклада R1a-M198 разделилась на субклады R1a-M417 и R1a-YP1051. Примерно 8000 лет назад носители субклада R1a-M417 мигрировали в северном направлении в сторону Кавказских гор, а носители субклада R1a-YP1051 в северно-западном направлении на территорию Анатолии. Носителей субклада R1a-YP1271 примерно 7500 лет назад мигрировала через Кавказские горы на территорию Северного Кавказа (рис.8).
в) Примерно в середине V-го тысячелетия до н.э. носители субклада R1a-YP1271 мигрировали с территории Северного Кавказа в Северное Причерноморье, а затем расселились практически по всей территории Европы. Носители субклада R1a-M417 в конце V-го тысячелетия до н.э. мигрировали на территорию Северного Кавказа, где разделились на три группы: первая группа мигрировала в западном направлении (носители субкладов R1a-Z283 и R1a-Z2464), вторая группа мигрировала в восточном направлении (носители субкладов R1a-Z93 (xZ94)), а третья группа мигрировала обратно в Закавказье (носители субклада R1a-Z95). Носители субклада R1a-YP1051 в середине IV-го тысячелетия до н.э. мигрировали через Анатолию на территорию Балканский полуостров (рис. 9).
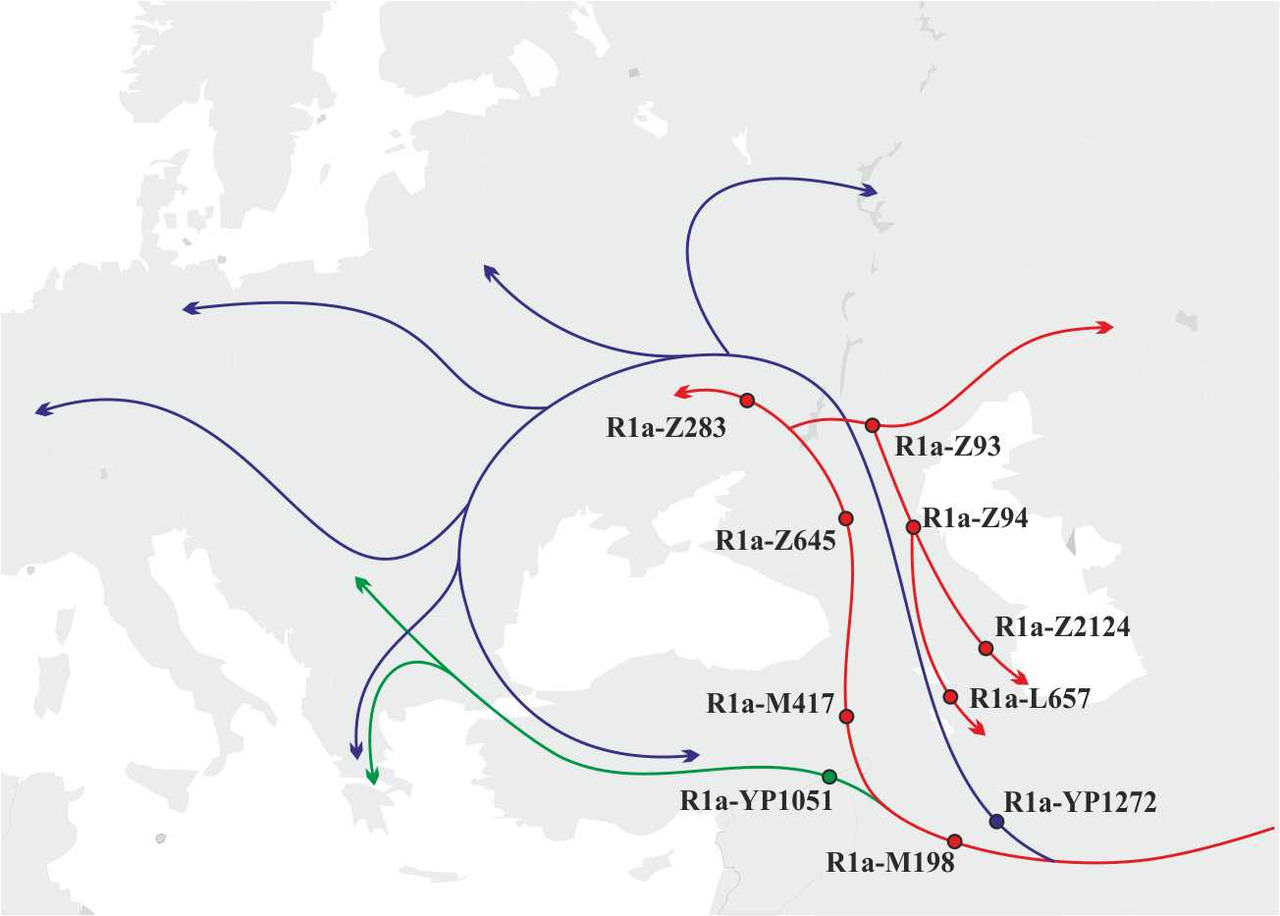
Носители субклада R1a-M417 оказалась самой успешной из этих трех групп носителей y-гаплогруппы R1a. Большинство носителей у-гаплогруппы R1a, живущих сегодня в Европе и Азии, являются потомками именно этой группы.
Носители y-гаплогруппы R1b так же, как и носители y-гаплогруппы R1a, мигрировали через Иранское нагорье в западном направлении (рис.10).
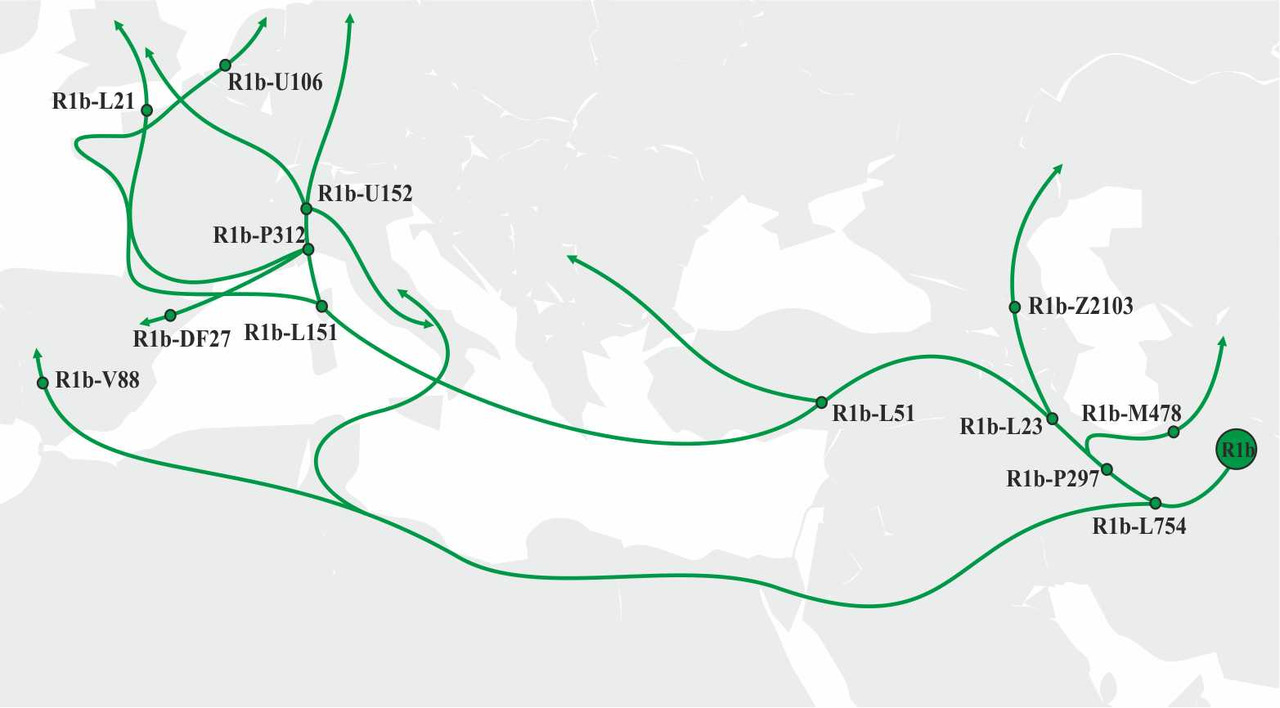
Миграция носителей y-гаплогруппы R1b на территорию Европы проходила по четырем основным маршрутам:
а) Носители субклада R1b-Z2103 мигрировали через Кавказские горы на территорию Северного Кавказа и далее в Поволжье [5];
б) Носители субклада R1b-L51 c территории Западной Анатолии мигрировали на Балканский полуостров и далее в центральную Европу;
в) Носители субклада R1b-L151 мигрировали c острова Корсика и расселились по территории Западной Европы;
г) Предки носителей субклада R1b-V88 мигрировали c территории Северной Африки на территорию Пиренейского полуострова.
Большинство носителей y-гаплогруппы R1b, проживающих сегодня на территории Европы, является носителями субклада R1b-L151. Носители остальных субкладов y-гаплогруппы R1b не были столь успешными и не смогли создать крупные популяции на территории Европы.
1.3. Соотнесение данных ДНК генеалогии с известными археологическими культурами
Исследование ДНК захоронений человеческих останков, безусловно, будет являться решающим аргументом при установлении популяции, проживавшей на определенной территории в определенный исторический период. Однако возможность определения временных границ миграций носителей интересующих нас y-гаплогрупп и их субкладов уже сейчас дает возможность сопоставить данные об этих миграциях с известными археологическими культурами. Рассмотрим на примере носителей y-гаплогруппы R1a новые возможности по установлению связи между известными археологическими культурами и миграциями носителей определенных y-гаплогрупп и их субкладов.
Примерно 9 000 тысяч лет назад носители субклада R1a-YP1271 мигрировали с территории Иранского нагорья в северном направлении в сторону Кавказских гор, а носители субклада R1a-M198 примерно в это же время мигрировала в северо-западном направлении на территорию Месопотамии, где произошло разделение этого субклада на субклады R1a-YP1051 и R1a-M417. Примерно в это же время на территории Месопотамии возникает одна из первых неолитических культур Ближнего Востока — хассунская культура (рис. 11).
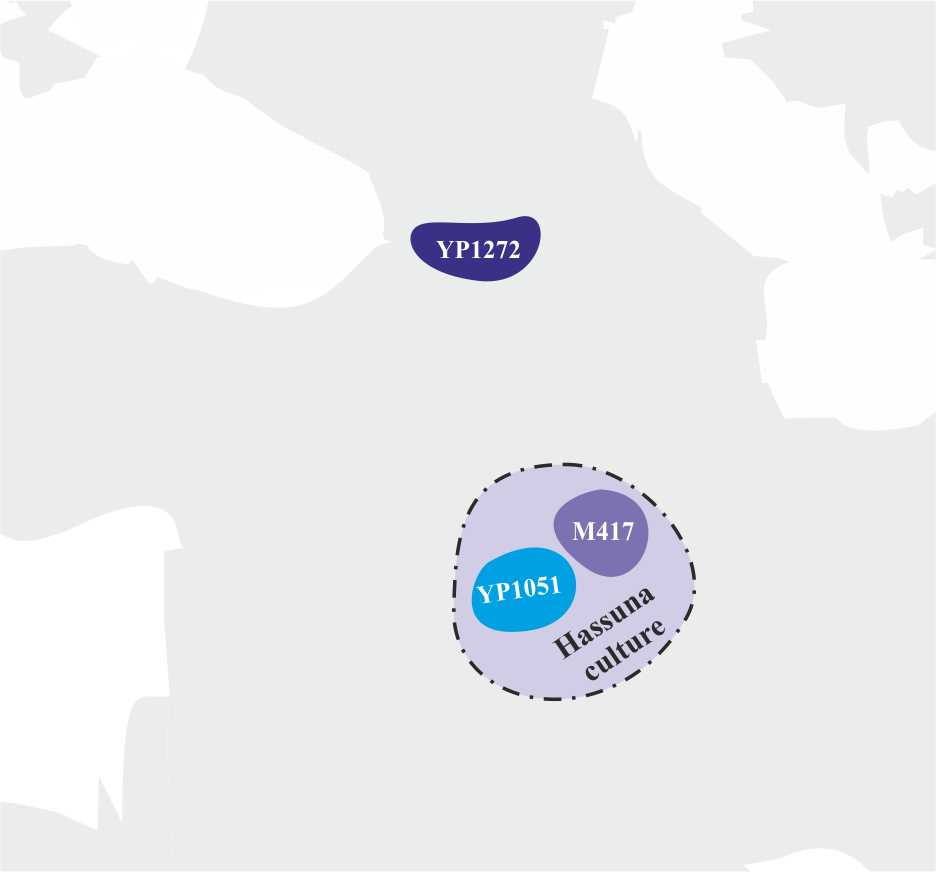
Совпадение времени возникновения на территории Месопотамии новой культуры с миграцией в этот регион носителей y-гаплогруппы R1a дает основание считать, что хассунская культура имеет прямое отношение к носителям субкладов R1a-YP1051 и R1a-M417.
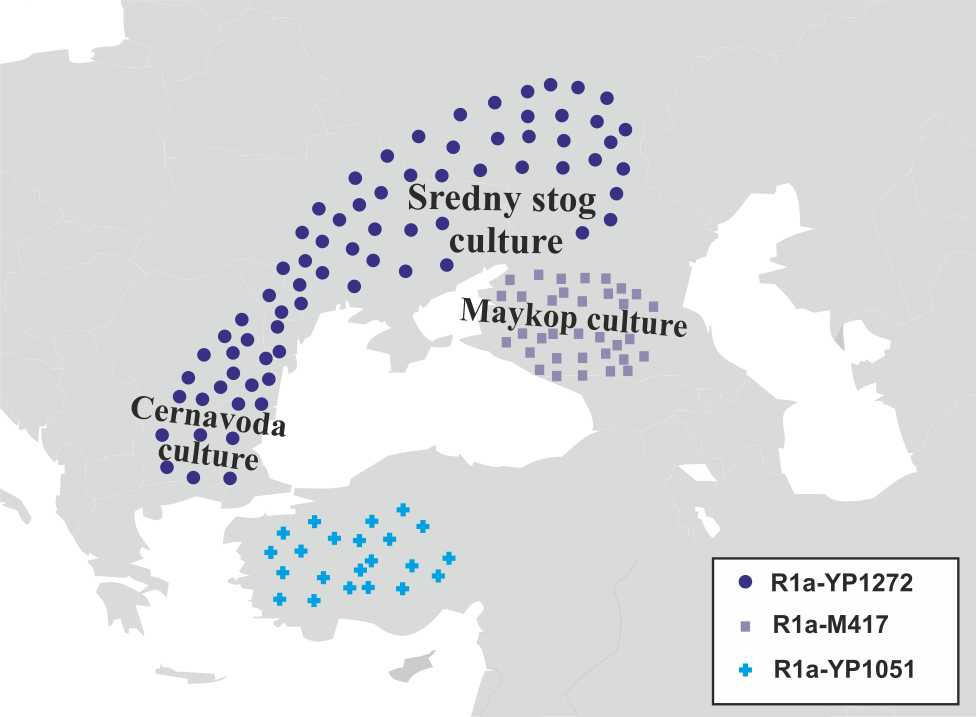
В середине VI-го тысячелетия до н.э. носители субклада R1a-YP1272 переходят через Кавказские горы и мигрируют далее в Северное Причерноморье, где в середине V-го тысячелетия до н.э. возникает среднестоговская культура. Носители субкладов R1a-M417 и R1a-YP1051 мигрировали с территории хассунской культуры далее на север на территорию халафской культуры, основным населением которой были носители y-гаплогрупп G2 и G1 (рис.12).
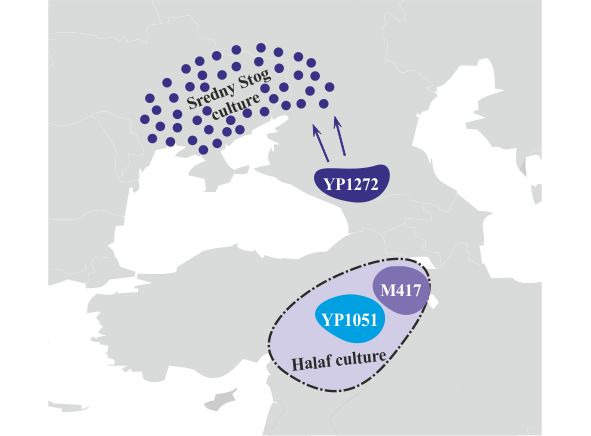
В конце V-го тысячелетия до н.э. носители субклада R1a-M417 мигрируют через Кавказские горы на территорию Северного Кавказа. В это же время на территории Северного Кавказа возникает майкопская культура. Совпадение времени возникновения майкопской культуры с миграцией носителей субклада R1a-M417 и связь этой культуры с культурами Северной Месопотамии дают основание предположить, что майкопская культура имеет прямое отношение к носителям субклада R1a-M417. Носители субклада R1a-YP1051 мигрируют в западном направлении через территорию Анатолии. В начале IV-го тысячелетия до н.э. носители субклада R1a-YP1272 начинают миграцию с территории Северного Причерноморья на территорию Балканского полуострова, где в это же время возникает культура Чернаводэ. Совпадение времени возникновения культуры Чернаводэ с миграцией носителей субклада R1a-YP1272 и ее связь со среднестоговской культурой дают основания предполагать, что культура Чернаводэ имеет прямое отношение к носителям субклада R1a- YP1272 (рис.13).
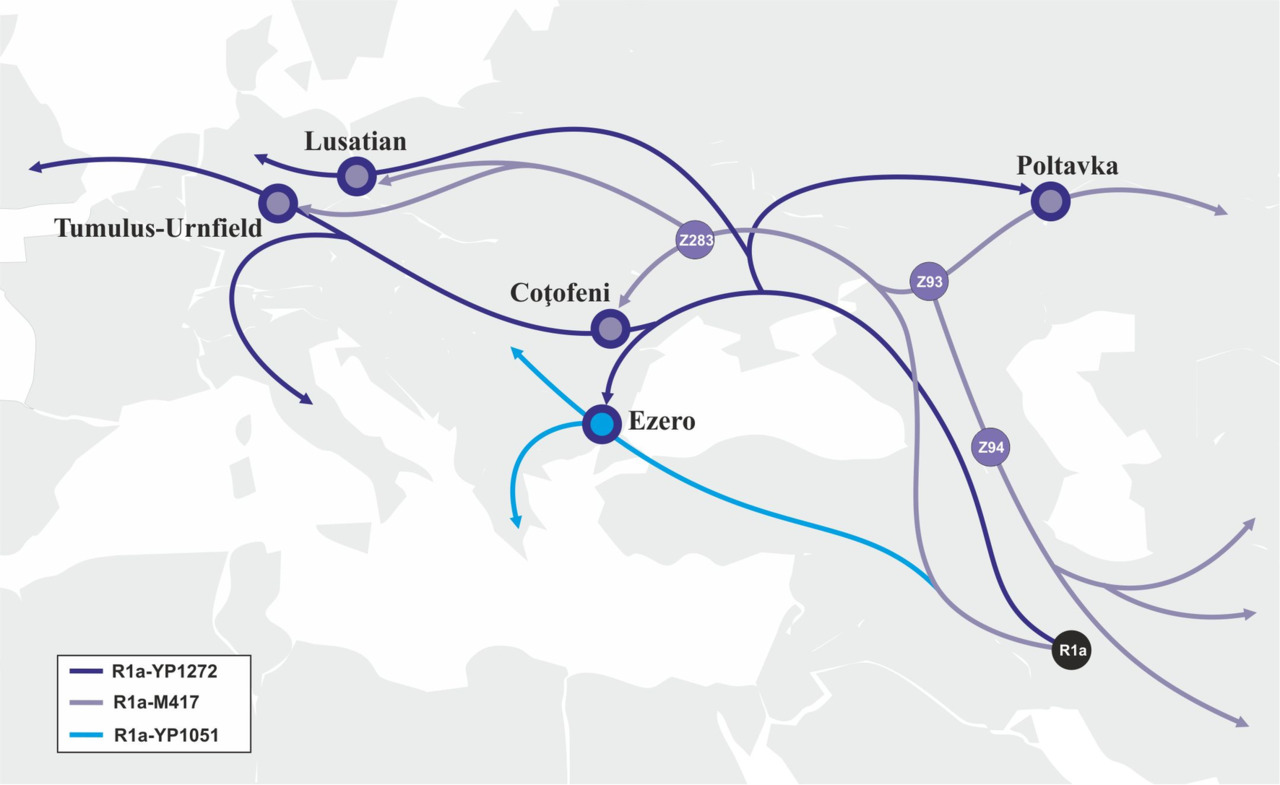
В середине IV-го тысячелетия до н.э. происходит миграция носителей субклада R1a-YP1272 с территории, прилегающей к побережью Черного моря, вглубь Европейского континента (баденская культура, западная часть ямной культуры). Часть носители субклада R1a-YP1051 мигрируют из Западной Анатолии на территорию Балканского полуострова. Примерно в это же время исчезает культура Чернаводэ и на ее месте возникает культура Езеро. Совпадение времени возникновения культуры Езеро с миграцией носителей субклада R1a-YP1051 и ее связь с анатолийскими культурами дают основания предполагать, что культура Езеро является результатом экспансии носителей субклада R1a-YP1051. Во второй половине VI тыс. до н.э. носители субклада R1a-M417 начинают проникать с территории майкопской культуры в Северное Причерноморье. В результате этих миграций происходит трансформация хвалынской и среднестоговской культур, а на их месте возникает ямная культура (рис.14).
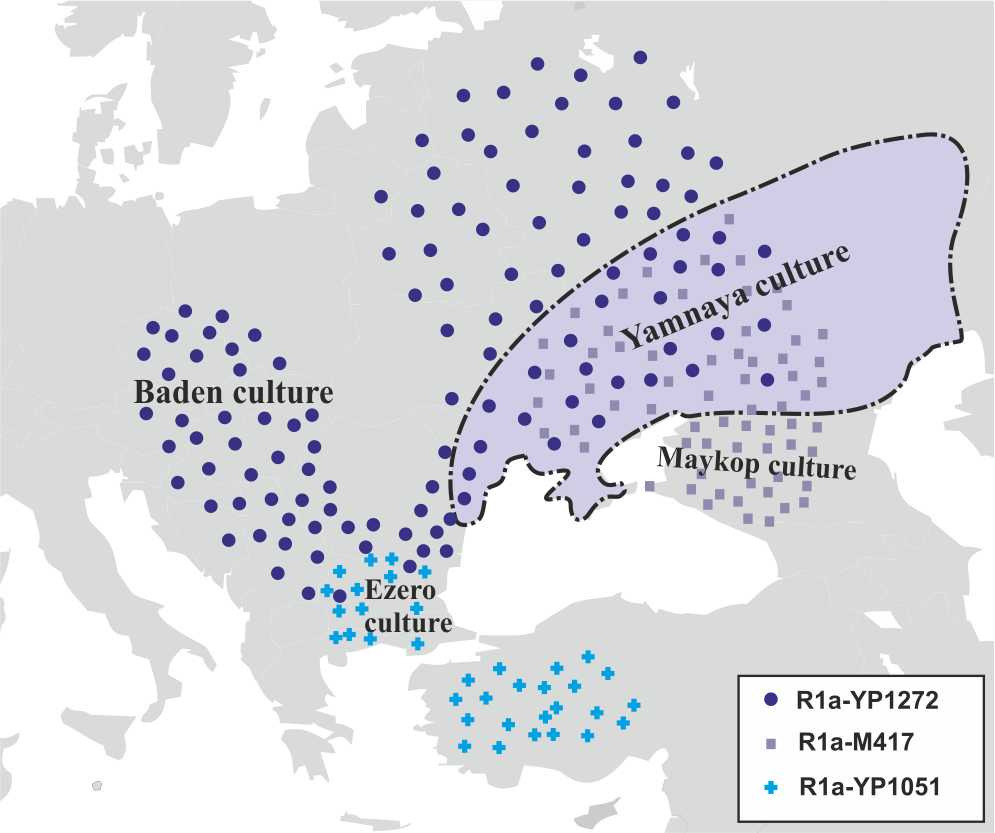
В начале III-го тысячелетия до н.э. происходит массовая миграция носителей субклада R1a-M417 с территории майкопской культуры в Северное Причерноморье. Эта экспансия приводит к выдавливанию с территории Северного Причерноморья население ямной культуры. На месте ямной культуры возникают катакомбная культура. Совпадение времени возникновения в Северном Причерноморье катакомбной культуры с миграцией носителей субклада R1a-M417 и ее связь с майкопской культурой дают основания предполагать, что катакомбная культура имеет прямое отношение к носителям субклада R1a-M417. Выдавленные с территории ямной культуры носители субкладов R1a-YP1272 и R1a-M417 мигрируют вглубь Европейского континента. В это же время на обширной территории Европейского континента возникает культура шнуровой керамики. Совпадение времени возникновения культуры шнуровой керамики с миграцией носителей субкладов R1a-YP1272 и R1a-M417 и ее связь с ямной культурой дают основания предполагать, что культура шнуровой керамики имеет прямое отношение к миграциям носителей субклада R1a-YP1272.
В середине III-го тысячелетия до н.э. начинается распад катакомбной культуры. Носители субклада R1a-L664 (возраст снипа ~4700 лет) мигрируют в Центральную Европу, а носители субклада R1a-Z284 (возраст снипа ~4500 лет) проникают на территорию Скандинавского полуострова. Носители субкладов R1a-L657 (возраст снипа ~4800 лет) и R1a-Z2124 (возраст снипа ~4800 лет) мигрируют в южном направлении в Закавказье. Примерно в это же время на территории Грузии возникает триалетская культура, имеющая корни в майкопской и катакомбной культурах. Носители субклада R1a-Z93 (xZ94) смещаются дальше на северо-восток в Среднее Поволжье. Примерно в это же время там возникает полтавкинская культура, имеющая корни в катакомбной культуре. Распределение основных субкладов y-гаплогруппы R1a к середине III тыс. до н.э. было примерно следующим (рис.15).
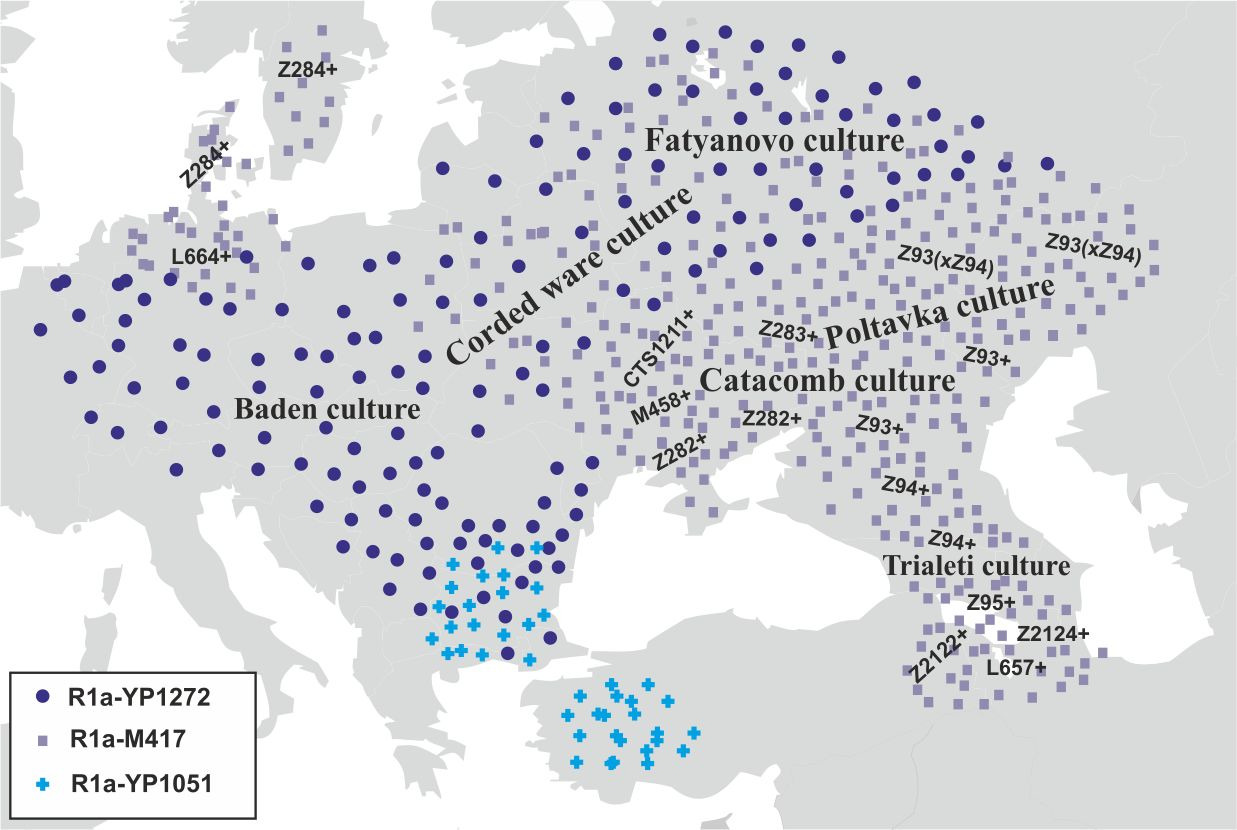
В начале II-го тысячелетия до н.э. носители субкладов R1a-Y35 (возраст снипа ~4300 лет) и R1a-YP343 (возраст снипа ~4300 лет) мигрировали на территорию Западной Украины и Восточной Польши, а носители субклада R1a-Z92 (возраст снипа ~4700 лет) заселили центр и северо-восток Украины. На территории, которую в начале II-го тысячелетия до н.э. заняли носители субкладов R1a-Y35, R1a-YP343 и R1a-Z92, возникает тшинецкая культура. Носители субклада R1a-M458 (возраст снипа ~4700 лет) мигрировали на юго-запад Украины, где в начале II-го тысячелетия до н.э. возникла комаровская культура. Часть носителей субкладов R1a-Z282* мигрирует на территорию Балканского полуострова. Носители субклада R1a-Z93 (xZ94) (возраст снипа ~5000 лет) в начале II-го тысячелетия до н.э. мигрируют далее на восток. Примерно в это же время на территории Казахстана и в Западной Сибири возникает андроновская культура. На месте катакомбной и полтавкинской культур формируется срубная культура. Носители субкладов R1a-L657 и R1a-Z2124 в начале II-го тысячелетия до н.э. начинают мигрировать с территории Закавказья на Ближний Восток, а далее Среднюю и Южную Азию (миграция ариев). Часть носителей субклада R1a-YP1272 мигрирует в Грецию и Анатолию. На территории России абашевская культура сменяет фатьяновскую. После всех этих миграций географическая локализация основных субкладов R1a была примерно следующей (рис.16).
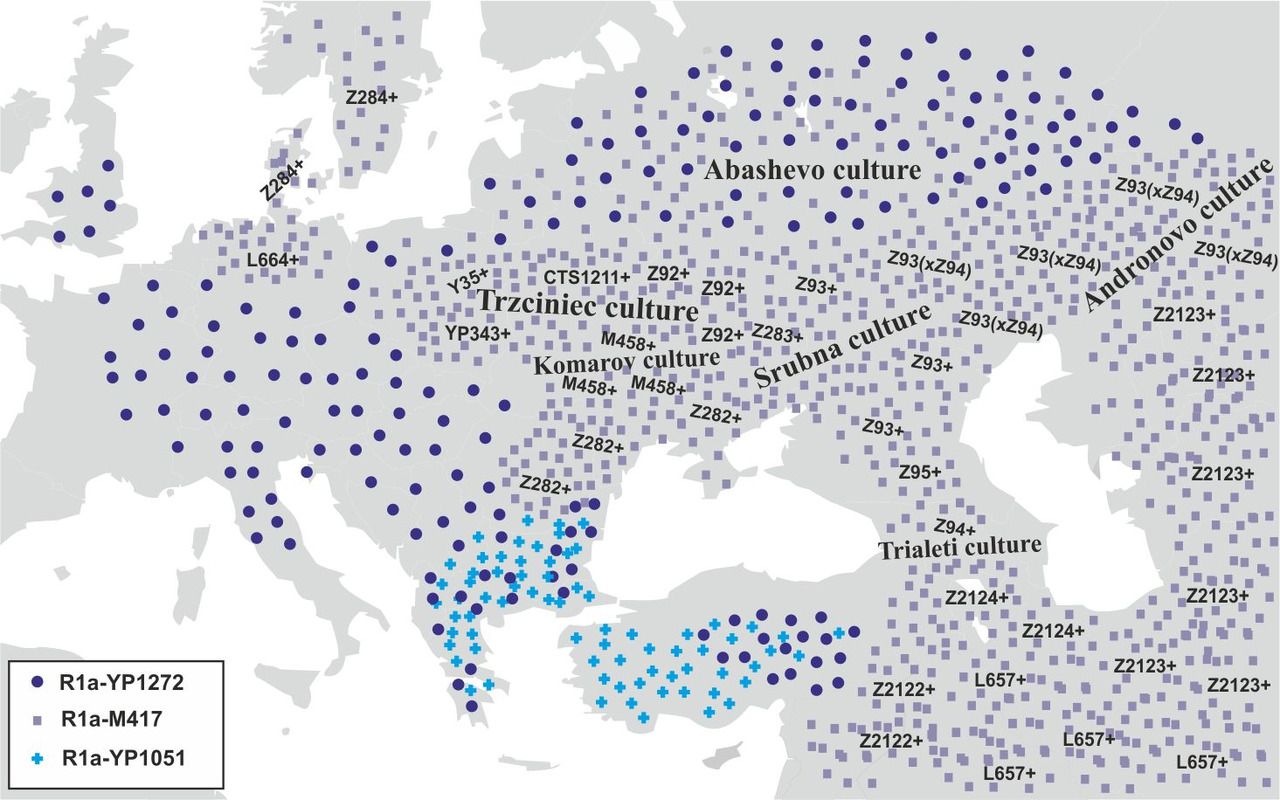
Примерно в 1600 году до н.э. носители субклада R1a-YP343 мигрировали с территории тшинецкой культуры далее на запад и вторглись на территорию унетицкой культуры. В результате этого вторжения унетицкая культура сменяется культурой курганных погребений. Носители субкладов R1a-Z282* мигрируют несколькими волнами с территории сегодняшней Румынии на юг Балканского полуострова и на территорию Анатолии. Примерно в это же время на территории Греции возникает микенская цивилизация. Экспансия носителей субкладов R1a-Z282* приводит к миграции населения Балканского полуострова на территорию Анатолии и Ближнего Востока (миграция «народов моря»). Часть населения срубной культуры (носителей субкладов R1a-Z93*) смещаются на запад на территорию Украины и Молдавии. Географическая локализация основных субкладов R1a к концу II-го тысячелетия до н.э. была примерно следующей (рис.17).
Основной этап расселения носителей y-гаплогруппы R1a закончился к концу II-го тыс. до н. э. Дальнейшие миграции носителей y-гаплогруппы R1a не были столь существенными и не привели к заметным изменениям структуры населения на территории Евразии.
Таким образом, в упрощенном виде миграции носителей y-гаплогруппы R1a можно схематично представить как последовательный выход трех крупных групп носителей данной y-гаплогруппы с территории Иранского нагорья, их разделение на территории Ближнего Востока в период раннего неолита, проникновение на территорию Европы и миграции в Центральную и Западную Европу в период бронзового и железного веков (рис.18).
Глава II. Теория сдвигов согласных звуков
2.1. Основная идея теории сдвигов согласных звуков
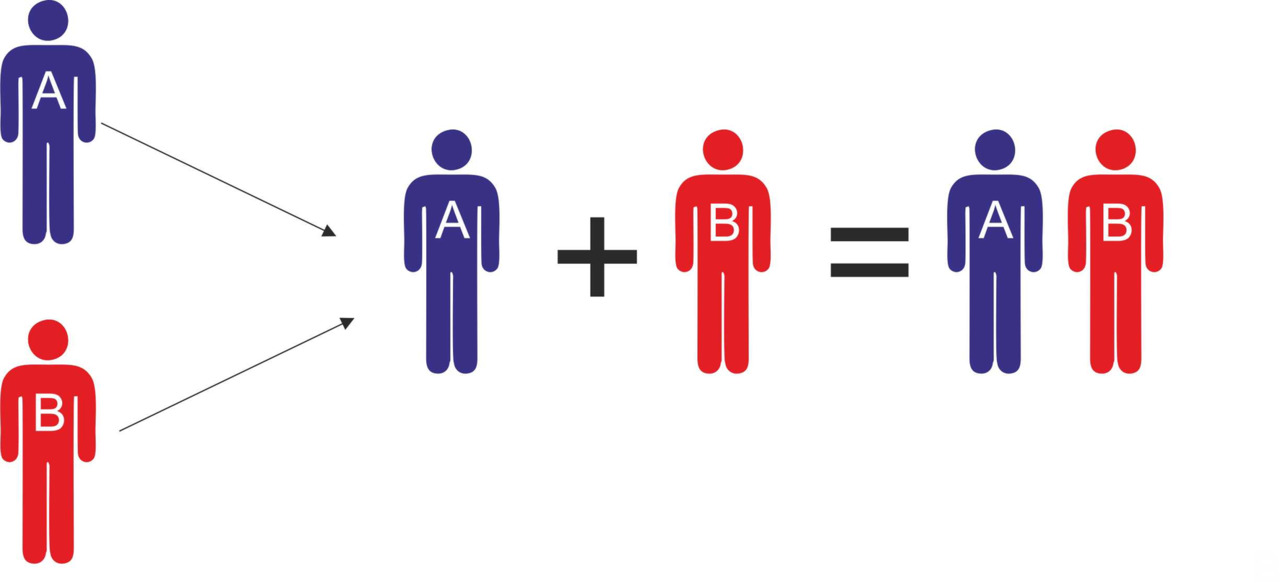
Использование информации о расселении носителей определенных y-гаплогрупп может существенно упростить моделирование процессов генезиса протоязыков. ДНК генеалогия дает возможность с достаточно высокой точностью определить популяции, которые могли участвовать в генезисе определенных протоязыков, а также установить временные и географические границы этих процессов. Информация о популяциях, принимавших участие в генезисе протоязыков, дает возможность установить связь между исходными языками популяций, принимавших участие в генезисе, и языками, возникшим в результате смешения этих популяций. Другими словами, если мы знаем, что язык AB является результатом смешения популяций A и B, мы можем сделать определенные предположения относительно исходных языков, на которых говорили популяции A и B до их смешения (рис.19).
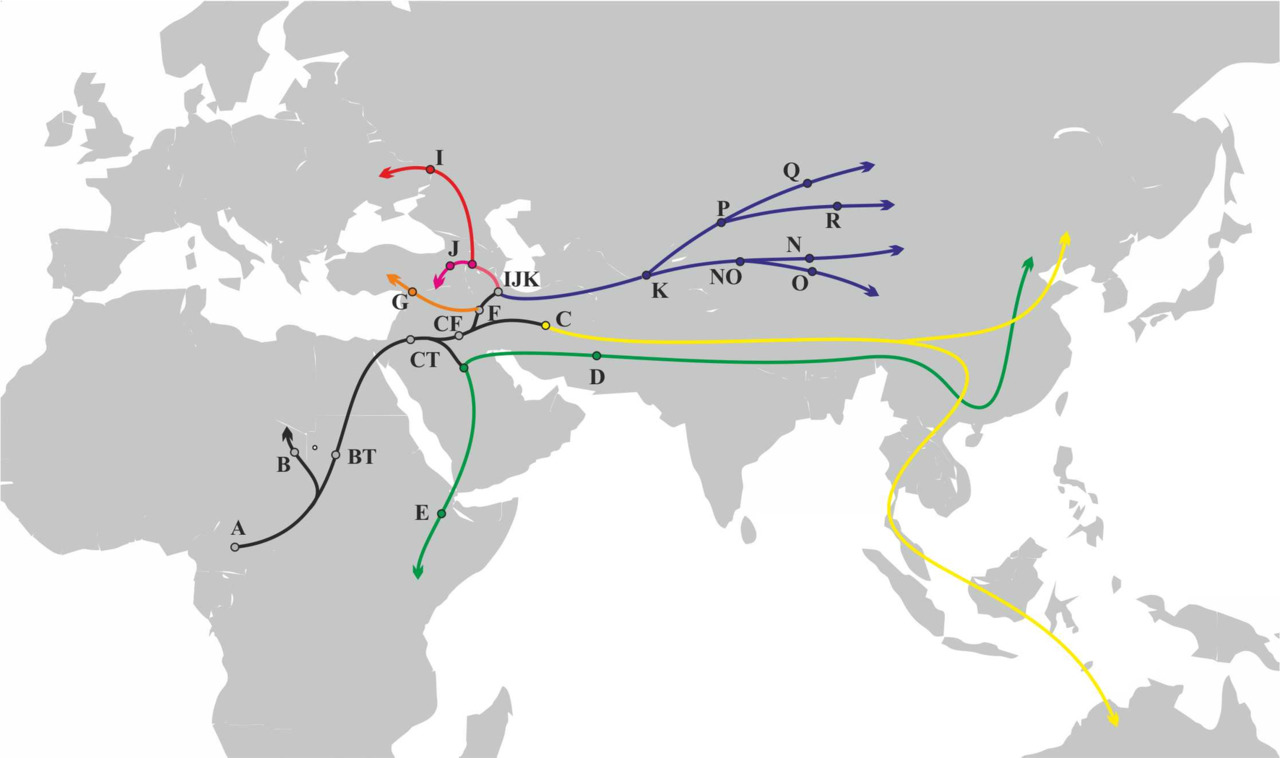
Если мы посмотрим на филогеническое дерево y-гаплогрупп, то увидим, что разделение на основные y-гаплогруппы произошло примерно ~65 000 — ~47000 лет назад. Информация о географической локализации носителей различных y-гаплогрупп дает возможность оценить вероятность контактов между носителями определенных y-гаплогрупп или об отсутствии таковых. Примерный маршрут миграции АСЧ после выхода с территории Африки может быть восстановлен на основании данных о распределении субкладов y-гаплогрупп в современных популяциях (рис.20).
Определив носителей y-гаплогрупп, которые могли иметь между собой контакты после разделения на соответствующие y-гаплогруппы, можно определить, какие лингвистические макроветви должны были возникнуть после первичного расселения АСЧ. С учетом географической локализации y-гаплогрупп и времени их разделения на отдельные y-гаплогруппы, все вновь возникшие в ходе миграций АСЧ в период с ~70 000 по ~45 000 гг. до н.э. языки можно разделить на пять макроветвей (рис.21).
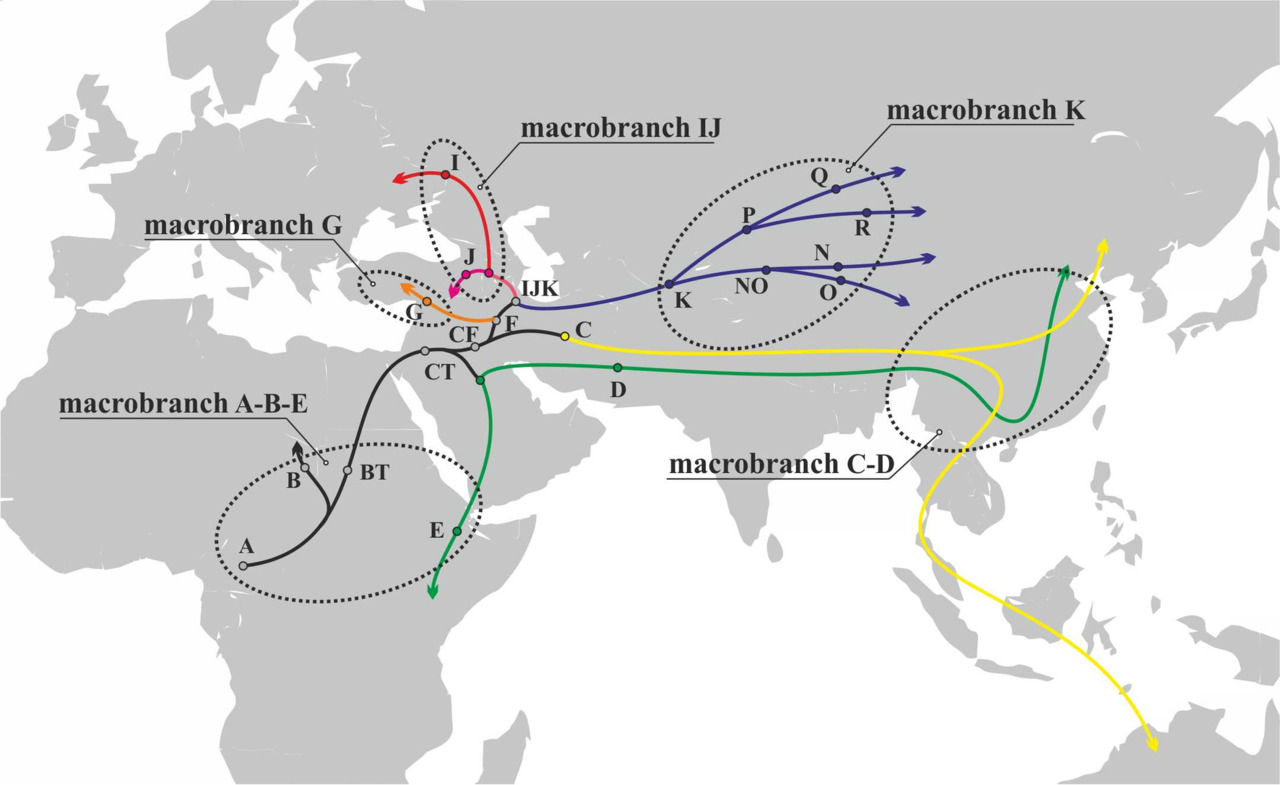
Таким образом, результатом разделения носителей различных y-гаплогрупп в ходе миграций АСЧ в период с ~70 000 по ~45 000 гг. до н.э. стало разделение исходного языка носителей y-гаплогруппы CT на следующие макроветви:
1) макроветвь A-B-E (включает в себя носителей y-гаплогрупп A, B и E);
2) макроветвь C-D (включает в себя носителей y-гаплогрупп C и D);
3) макроветвь G (носители y-гаплогруппы G);
4) макроветвь IJ (включает в себя носителей y-гаплогруппI и J);
5) макроветвь K (включает в себя носителей y-гаплогруппO, N, Q и R);
Схематически разделение исходного языка носителей y-гаплогруппы CT на макроветви в период с ~70 000 по ~45 000 гг. до н.э. может быть представлено следующим образом (рис.22).
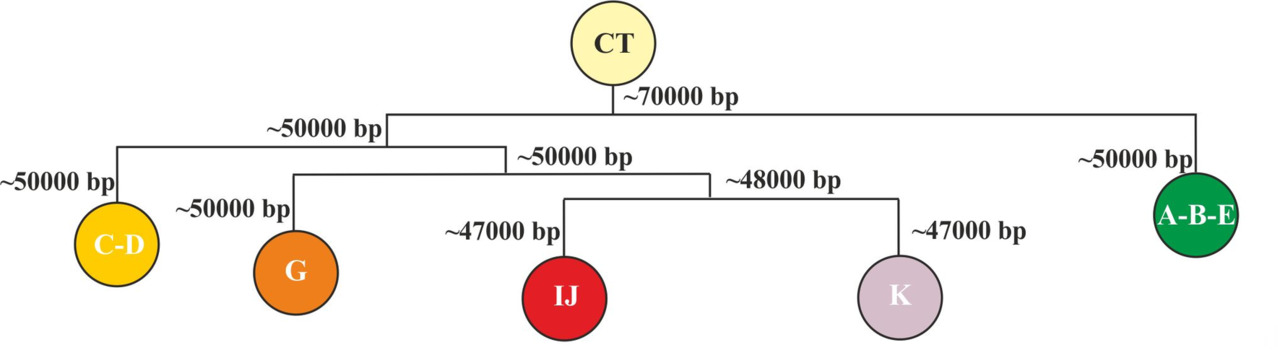
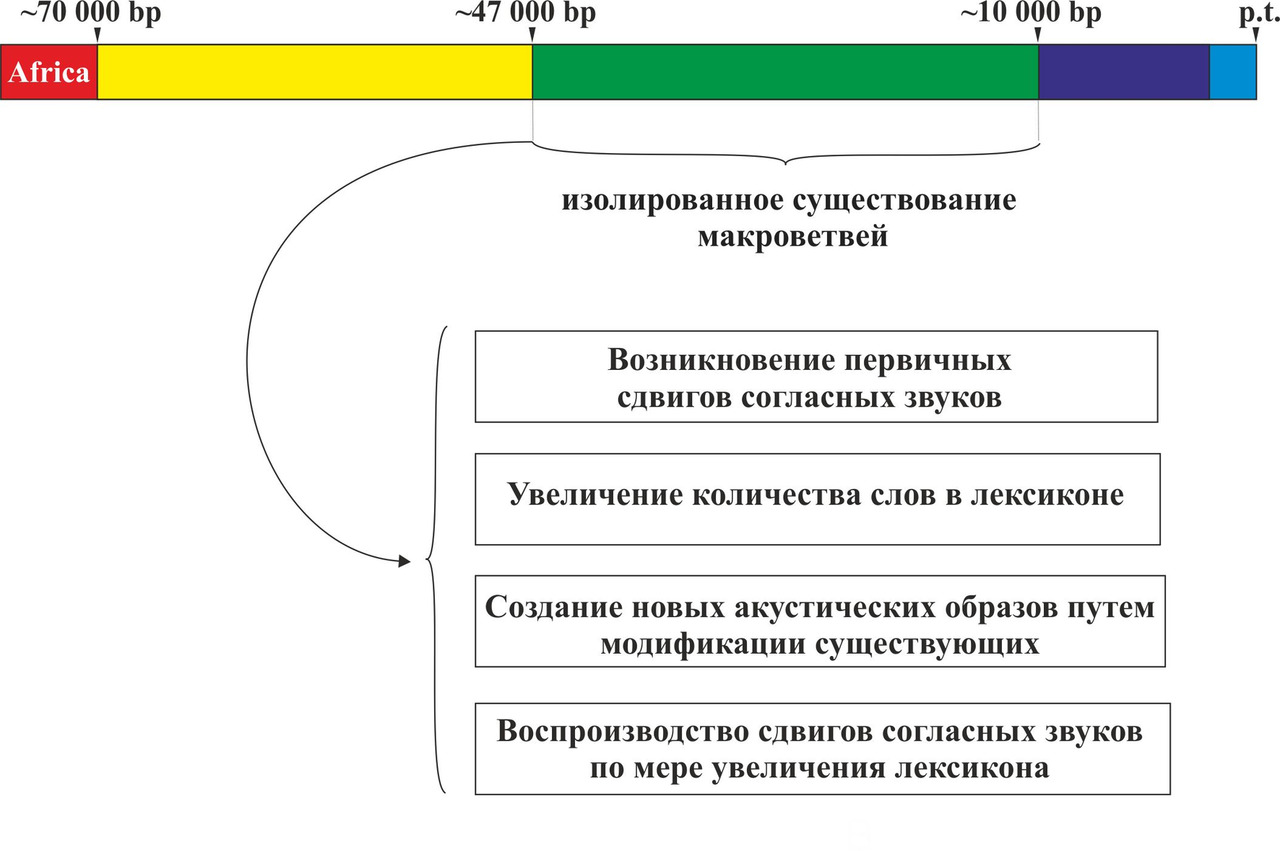
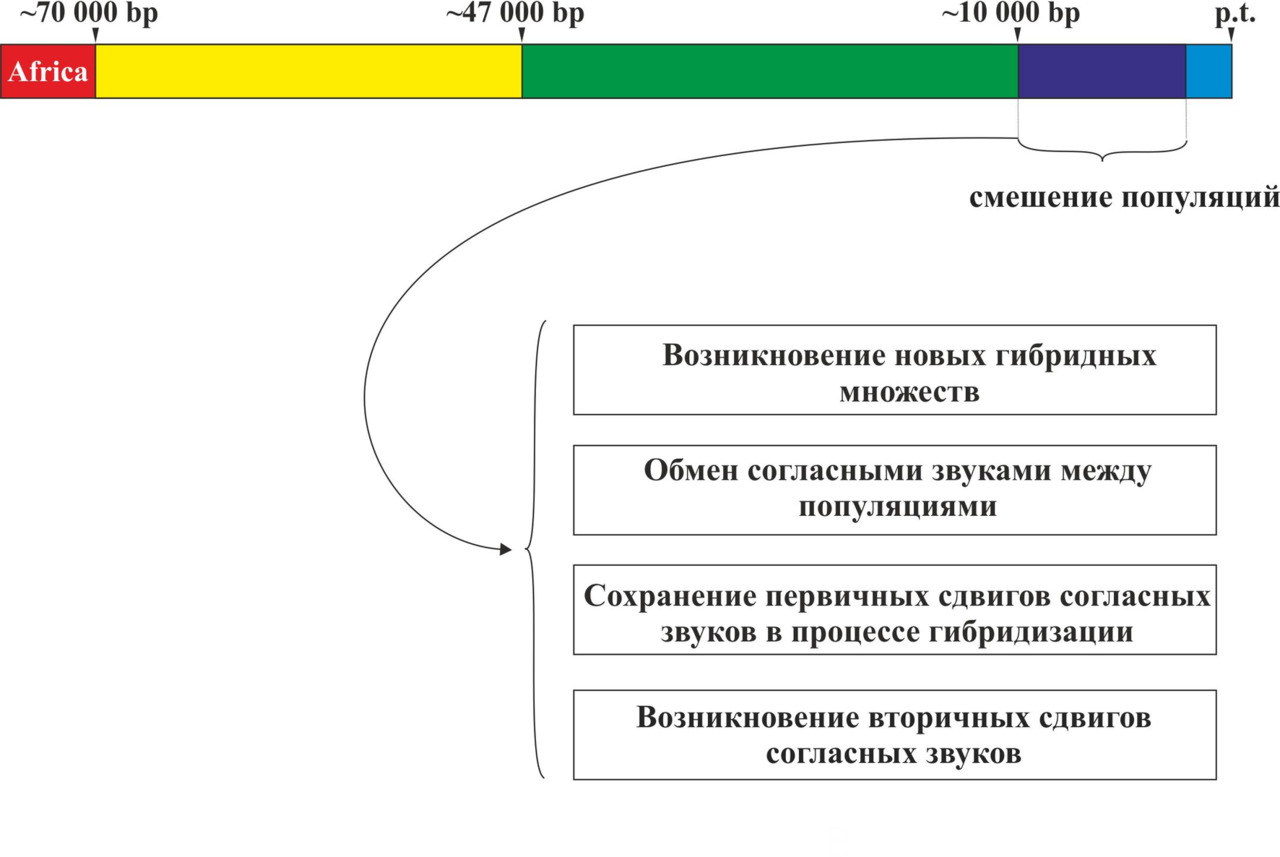
Таким образом, все современные языки являются прямыми потомками данных пяти макроветвей или являются результатом их смешения. При этом развитие языков после выхода носителей y-гаплогруппы CT с территории Африки можно разбить на три основных этапа: разделение на основные y-гаплогруппы и формирование макроветвей (период с ~70 000 по ~45 000 гг. до н.э.), изолированное существование макроветвей (период с ~45 000 по ~8 000 гг. до н.э.), смешение популяций, говоривших на языках, относящихся к разным макроветвям (период с ~8 000 по ~2 000 гг. до н.э.) (рис. 23).
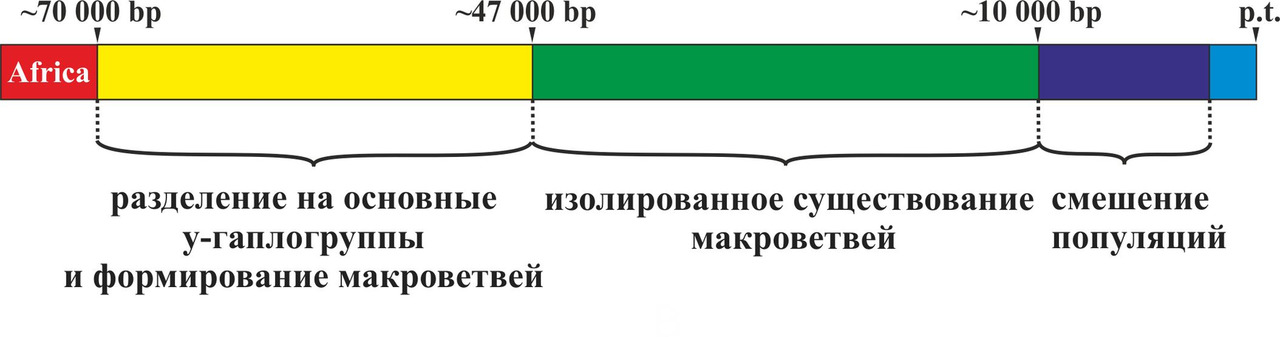
Основной идеей теории сдвигов согласных звуков является предположение о возникновении в изолированных друг от друга популяциях первобытных людей (период с ~45 000 по 8 000 гг. до н.э.) сдвигов согласных звуков (первичные сдвиги), которые были специфичными для каждой макроветви. В ходе процесса смешения популяций (период с ~8 000 по 2 000 гг. до н.э.) произошли вторичные сдвиги согласных звуков. При этом вторичные сдвиги согласных звуков должны были зависеть от первичных сдвигов, возникших в соответствующих макроветвях в период изолированного существования популяций. Рассмотрим более подробно причины возникновения первичных и вторичных сдвигов согласных звуков.
2.2. Причины первичных и вторичных сдвигов согласных звуков
Будет обоснованным предположить, что количество слов в лексиконе первобытных людей, вышедших с территории Африки (уровень y-гаплогруппы CT), не превышало нескольких десятков. Соответственно, первоначальное количество согласных звуков в языке первобытных людей на уровне y-гаплогруппы CT, скорее всего, не превышало пяти (множество согласных звуков С (CT)). Во время периода изолированного существования в каждой макроветви по мере роста количества слов в лексиконе должны были появиться новые согласные звуки для создания необходимого количества акустических образов. При этом вновь появившиеся согласные звуки, скорее всего, были специфичными для каждой из макроветвей (рис.24).
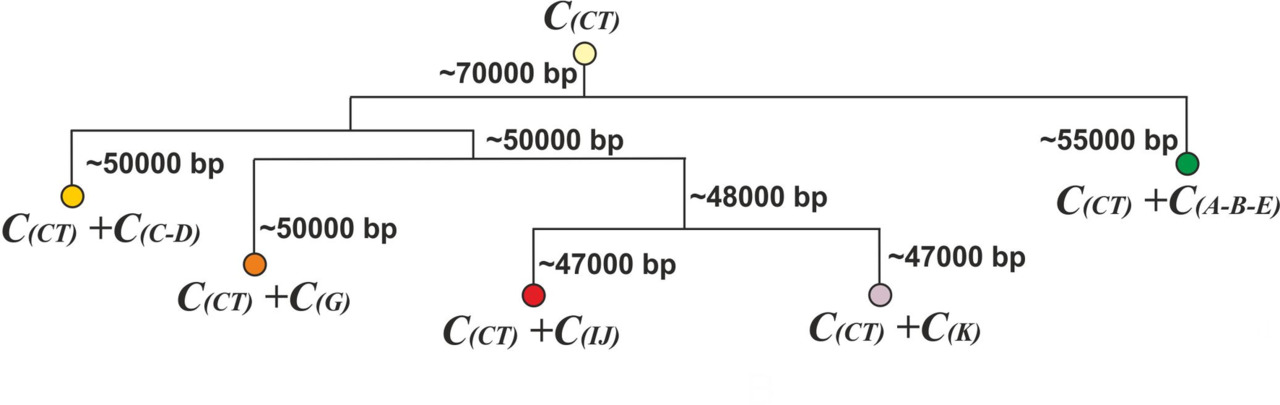
Таким образом, в результате первичных сдвигов согласных звуков в каждой из макроветвей к исходному множеству согласных звуков С (CT) прибавились специфичные для каждой макроветви согласные звуки: для макроветви A-B-E — множество С (A-B-E), для макроветви C-D — множество С (C-D), для макроветви G — множество С (G), для макроветви IJ — множество С (IJ), для макроветви K — множество С (K). При этом будет разумным предположить, что для каждой макроветви существовала своя характерная локализация новых согласных звуков. Например, если у какой-то макроветви первым новым согласным звуком оказался зубной согласный звук, то следующим новым согласным звуком тоже, скорее всего, был зубной звук, а если первым оказался губной согласный звук, то следующим тоже, скорее всего, был губной. Другими словами, создание новых согласных звуков должно было происходить путем вариации уже имевшихся согласных звуков. В результате у каждой макроветви в период изолированного существования должны были возникнуть свои специфичные наборы согласных звуков. Соответственно, в популяциях, принадлежавших к разным макроветвям, в период их изолированного существования должны были появиться новые акустические образы, возникшие после первичных сдвигов согласных звуков.
После периода длительной изоляции наступает период смешения популяций. Данный период связан с неолитической революцией и переходом от охоты и собирательства к земледелию и скотоводству. Увеличение численности популяций первобытных людей приводит к возникновению крупных поселений и даже протогорода. В результате всех этих процессов происходит смешение различных популяций древних людей и, как следствие, возникают гибридные языки. Гибридизация популяций должна была приводить к возникновению вторичных сдвигов согласных звуков. При этом вторичные сдвиги согласных звуков в гибридных популяциях должны были зависеть от первичных сдвигов согласных звуков в исходных популяциях. Информация о структуре ДНК современных популяций дает возможность сопоставить вторичные сдвиги согласных звуков в интересующих нас языках с процессами гибридизации носителей определенных y-гаплогрупп (рис. 25).
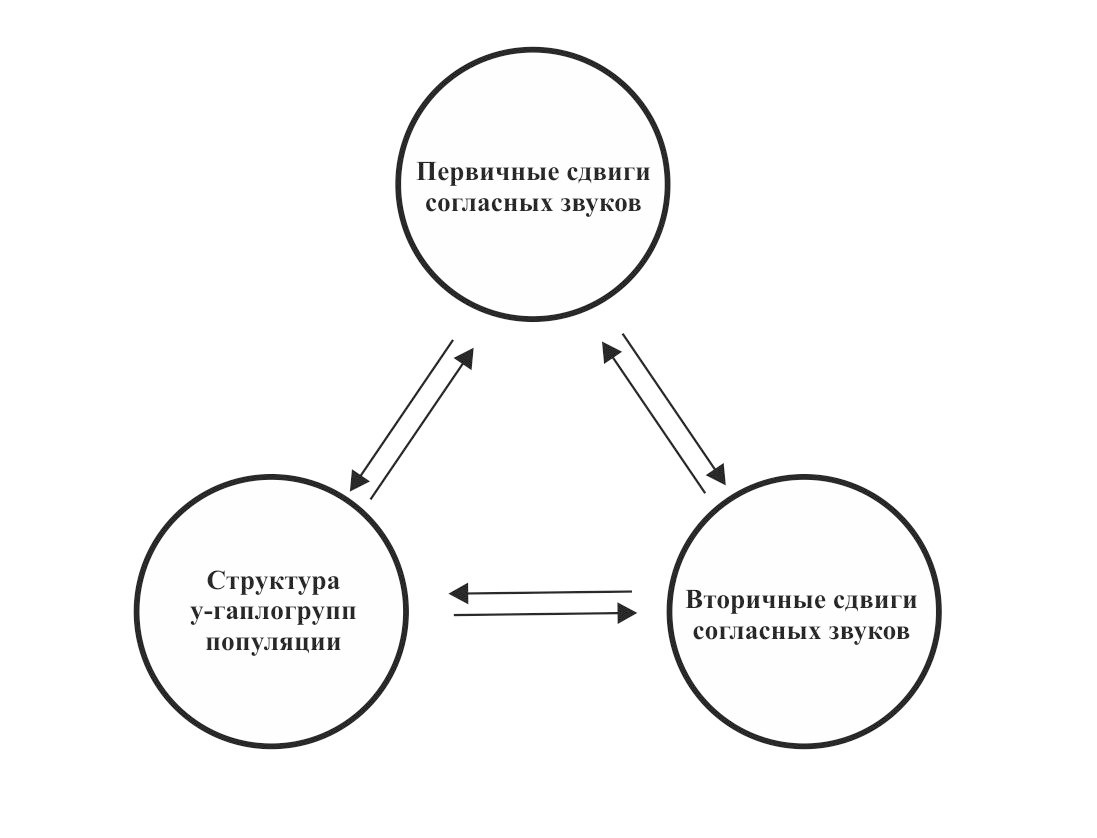
Таким образом, основной идеей теории сдвигов согласных звуков является предположение о существовании зависимости между сдвигами согласных звуков (первичными и вторичными) в языках гибридных популяций и структурой y-гаплогрупп этих популяций.
2.3. Популяции, принимавшие участие в генезисе индоевропейских языков
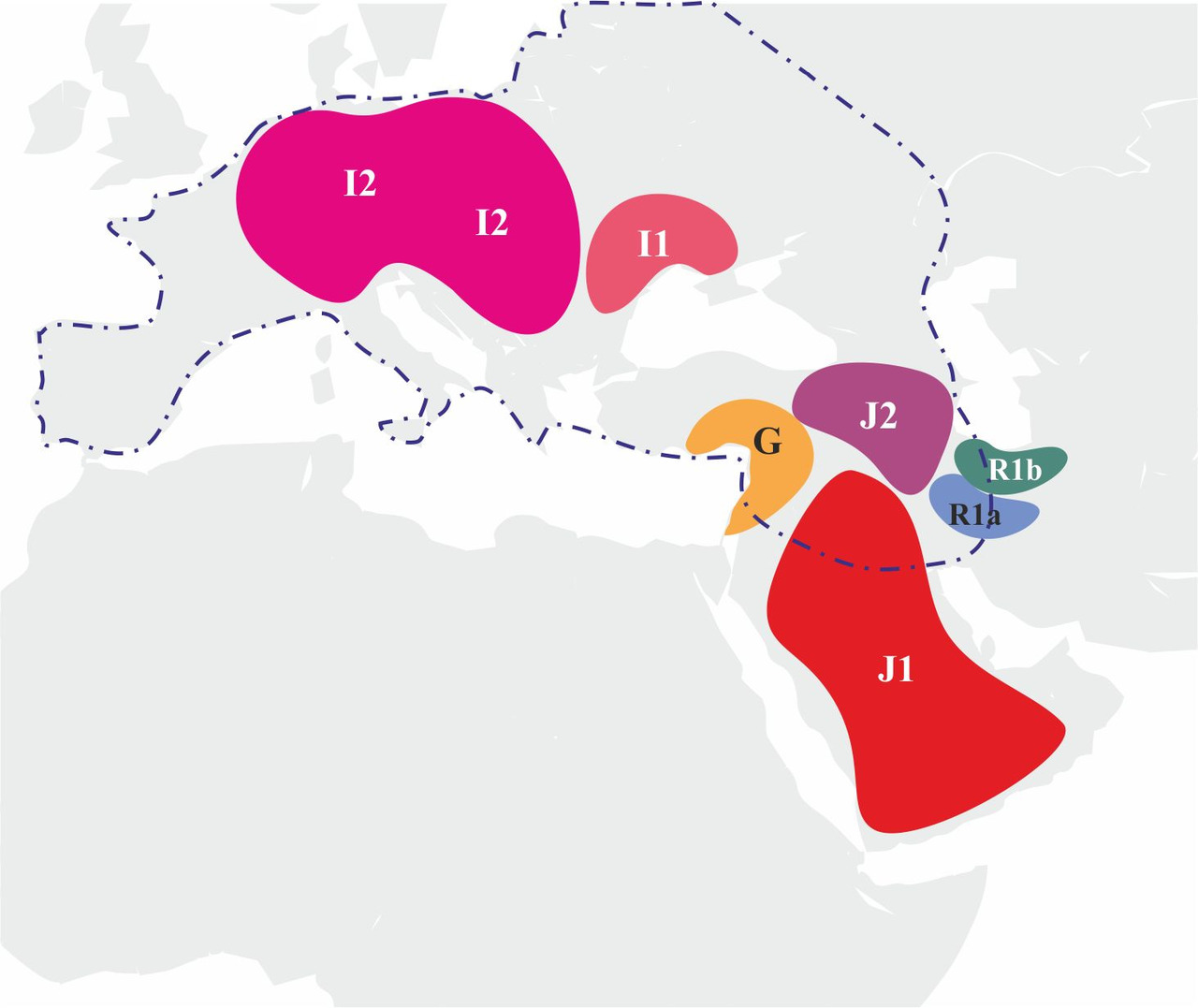
Для создания модели генезиса индоевропейских языков на территории Евразии необходимо определить круг популяций, которые принимали непосредственное участие в генезисе и распространении индоевропейских языков. На сегодняшний день основным источником информации о древних популяциях, принимавших участие в генезисе индоевропейских языков, являются результаты исследований ДНК останков древних захоронений различных археологических культур, существовавших на территории Евразии в период неолита и бронзового века. Информации о результатах таких исследований на сегодняшний день не так много, но, тем не менее, уже полученные на настоящий момент результаты исследований дают возможность сделать обоснованные предположения о носителях y-гаплогрупп, принимавших участие в генезисе индоевропейских языков на территории Европы. Согласно результатам исследований ДНК древних останков, которые были опубликованы в статьях «Genome flux and stasis in a five millennium transect of European prehistory» Christine Gamba et al от 21.10.2014 и «Ancient DNA reveals male diffusion through the Neolithic Mediterranean route» Marie Lacan et al. от 14.06.2011, в популяциях эпохи неолита и раннего бронзового века на территории Европы преобладали носители y-гаплогрупп I, G2 и R1a (рис.26).
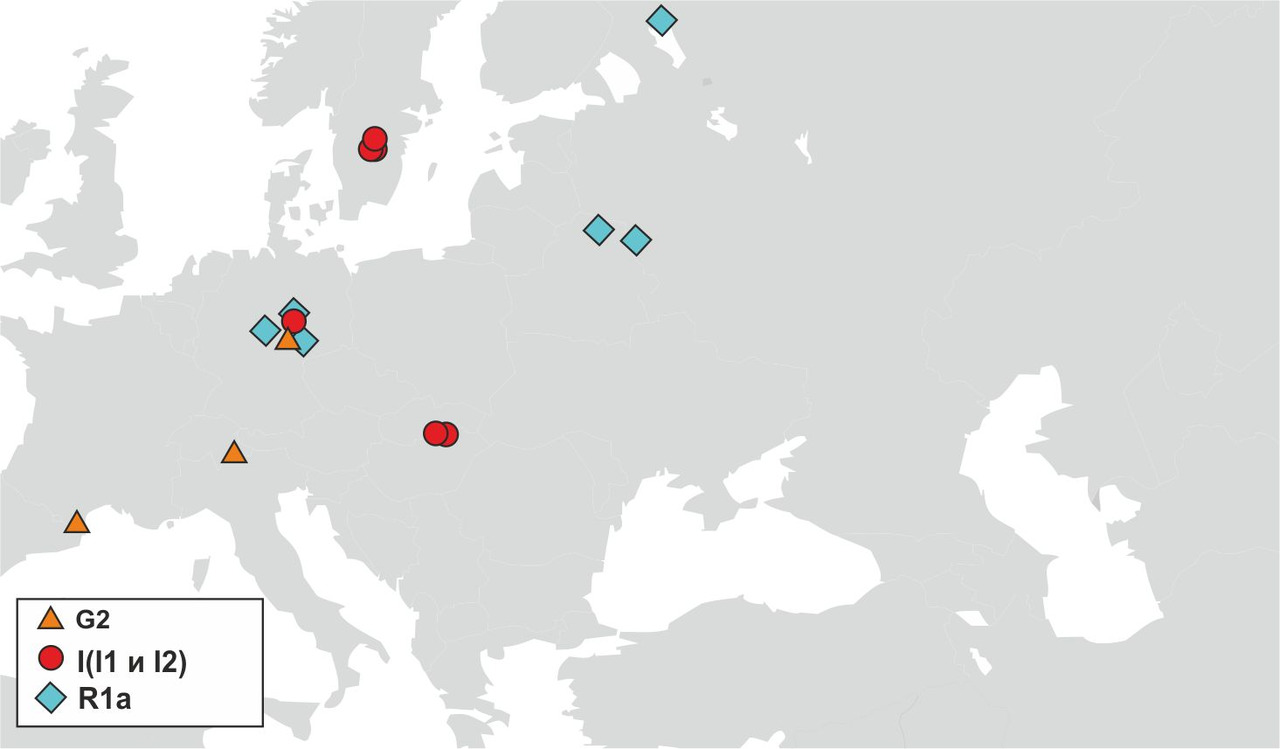
Исследований ДНК останков древних захоронений эпохи неолита и раннего бронзового века на территории Азии пока не проводилось. Тем не менее, данные о распределении y-гаплогрупп в современных популяциях, проживающих в Азии, дают возможность сделать обоснованные предположения о распределении основных y-гаплогрупп до начала массовых миграций периода неолита (рис.27).
Результаты исследований ДНК останков древних захоронений эпохи неолита и раннего бронзового века на территории Европы дают возможность ограничить круг популяций, принимавших участие в генезисе индоевропейских языков, а также определить маршруты миграций этих популяций (рис.28).
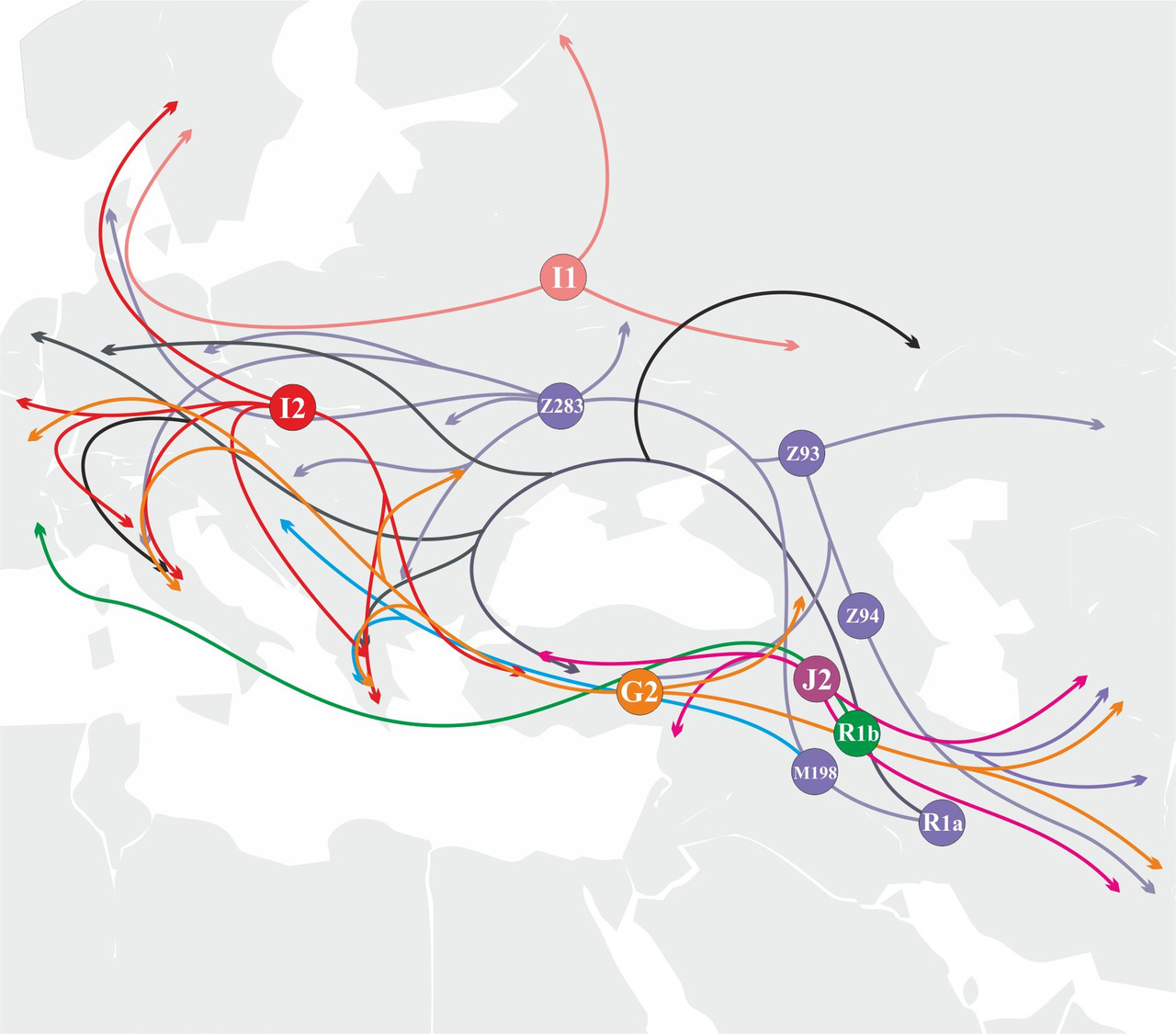
Очевидно, к основным популяциям, принимавшим участие в генезисе индоевропейских языков, необходимо отнести y-гаплогруппы I1, I2, J2, R1a, R1b и G2. Тот факт, что популяции, принимавшие участие в генезисе индоевропейских языков, относятся всего к трем разным макроветвям (G, IJ и K), существенно упрощает задачу определения первичных сдвигов согласных звуков специфичных для каждой из этих макроветвей. Первичные сдвиги согласных звуков в исходных популяциях можно определить по доминирующим сдвигам в языках гибридных популяций: если в гибридной популяции преобладали носители y-гаплогруппы I (макроветвь IJ), то в гибридном языке будет преобладать сдвиг согласных звуков специфичный для макроветви IJ, если преобладали носители y-гаплогруппы R1a (макроветвь K), то в гибридном языке будет преобладать сдвиг согласных звуков специфичный для макроветви K, а если преобладали носители y-гаплогруппы G2 (макроветвь G), то в гибридном языке будет преобладать сдвиг согласных звуков специфичный для макроветви G. Таким образом, на основании данных о степени участия определенных популяций в генезисе индоевропейских языков можно определить первичные сдвиги в исходных популяциях и в соответствующих им макроветвях.
С учетом известных на сегодняшний день миграций древних популяций участие носителей основных y-гаплогрупп в генезисе определенных ветвей индоевропейских языков на территории Евразии в период с IV-го по I-ое тысячелетия до н.э. может быть схематично представлено следующим образом (рис.29).
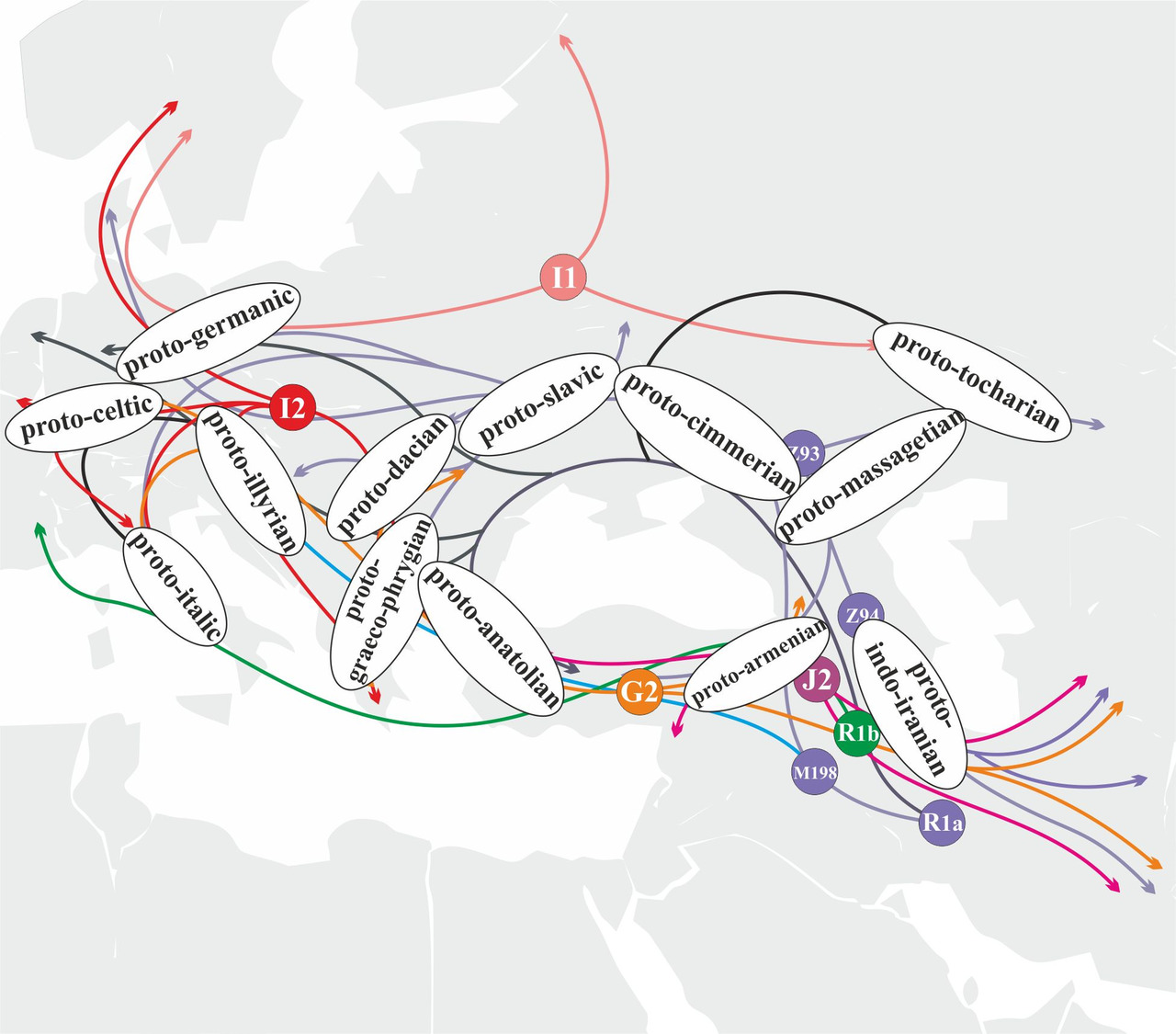
Анализа маршрутов миграций носителей различных y-гаплогрупп и их субкладов дает возможность достаточно точно оценить вероятность их участия в генезисе определенных ветвей индоевропейских языков. Например, носители y-гаплогруппы R1a принимали участие в формировании абсолютно всех ветвей индоевропейских языков. Из трех основных евразийских ветвей y-гаплогруппы R1a главную роль в генезисе и распространении индоевропейских языков играли носители субклада R1a-M417 (рис.30).
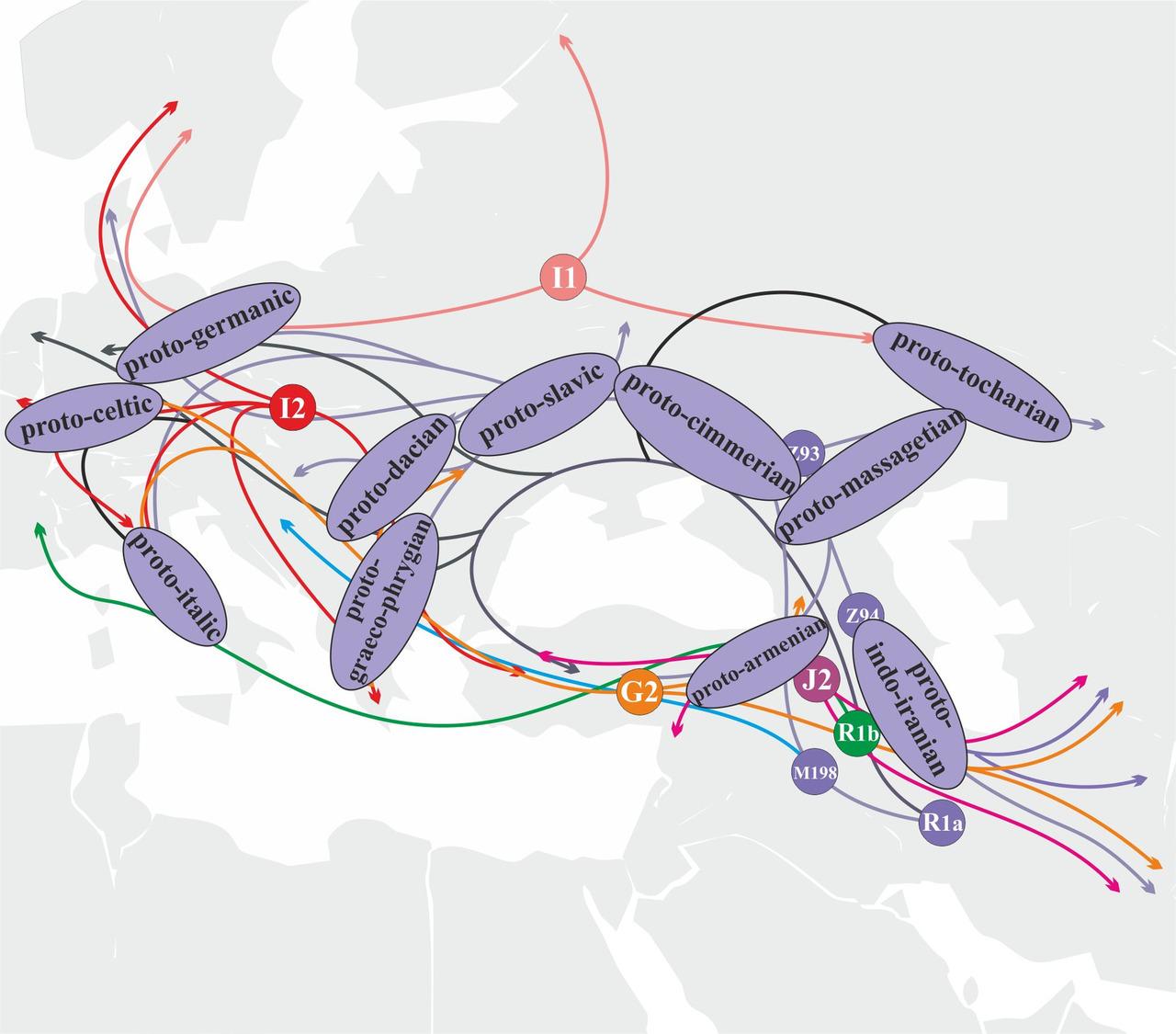
Носители субклада R1a-M417 приняли активное участие в генезисе большинства ветвей индоевропейских языков.
Носители субклада R1a-YP1272 прибыли на территорию Европы в первой волне мигрантов с Ближнего Востока и принимали активное участие в формировании индоевропейских языков Центральной и Южной Европы (рис.31).
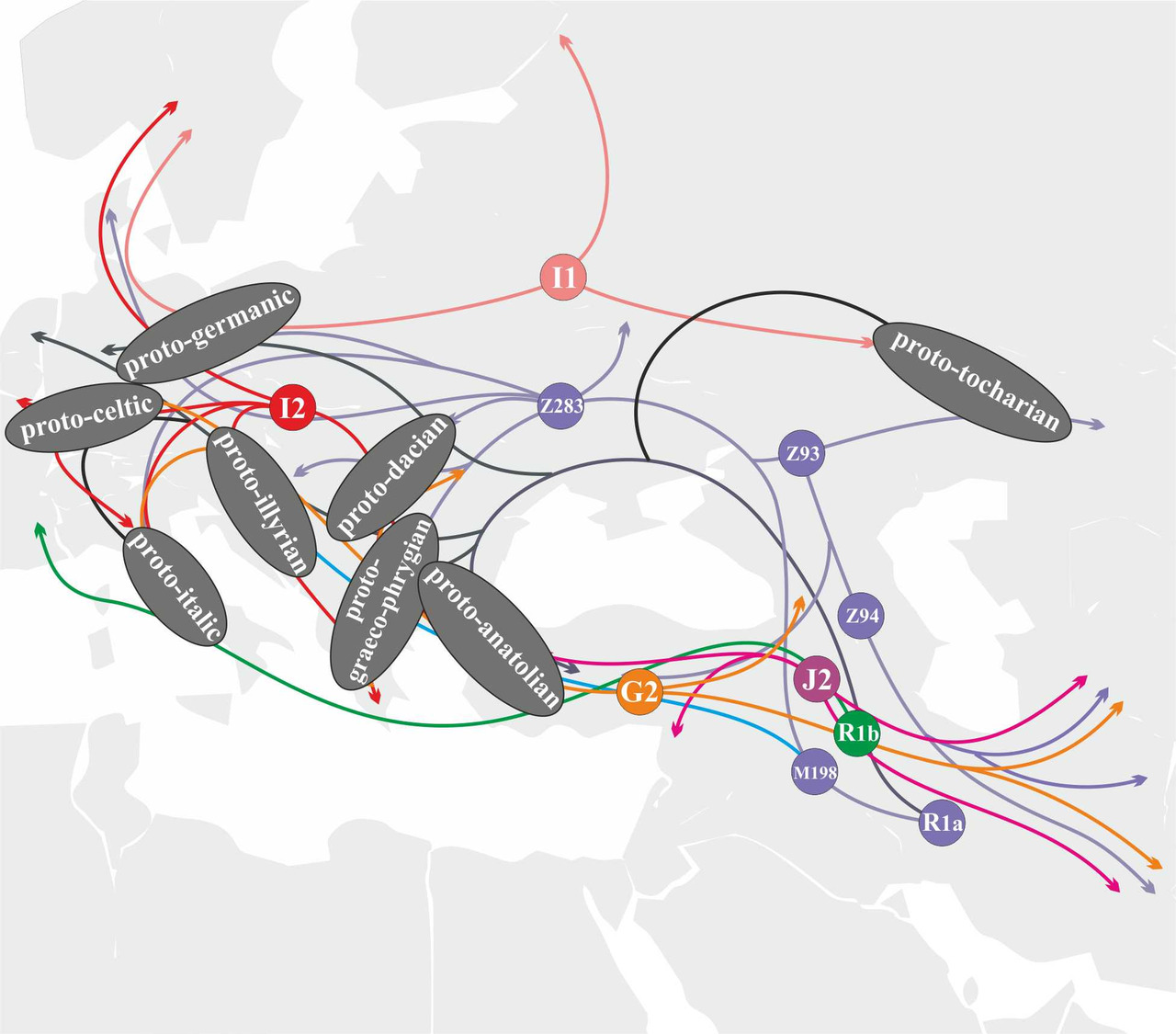
Носители субклада R1a-YP1051 мигрировали на территорию Европы из Западной Анатолии и принимали активное участие в формировании протоанатолийских и протогреко-фригийского языков (рис.32).
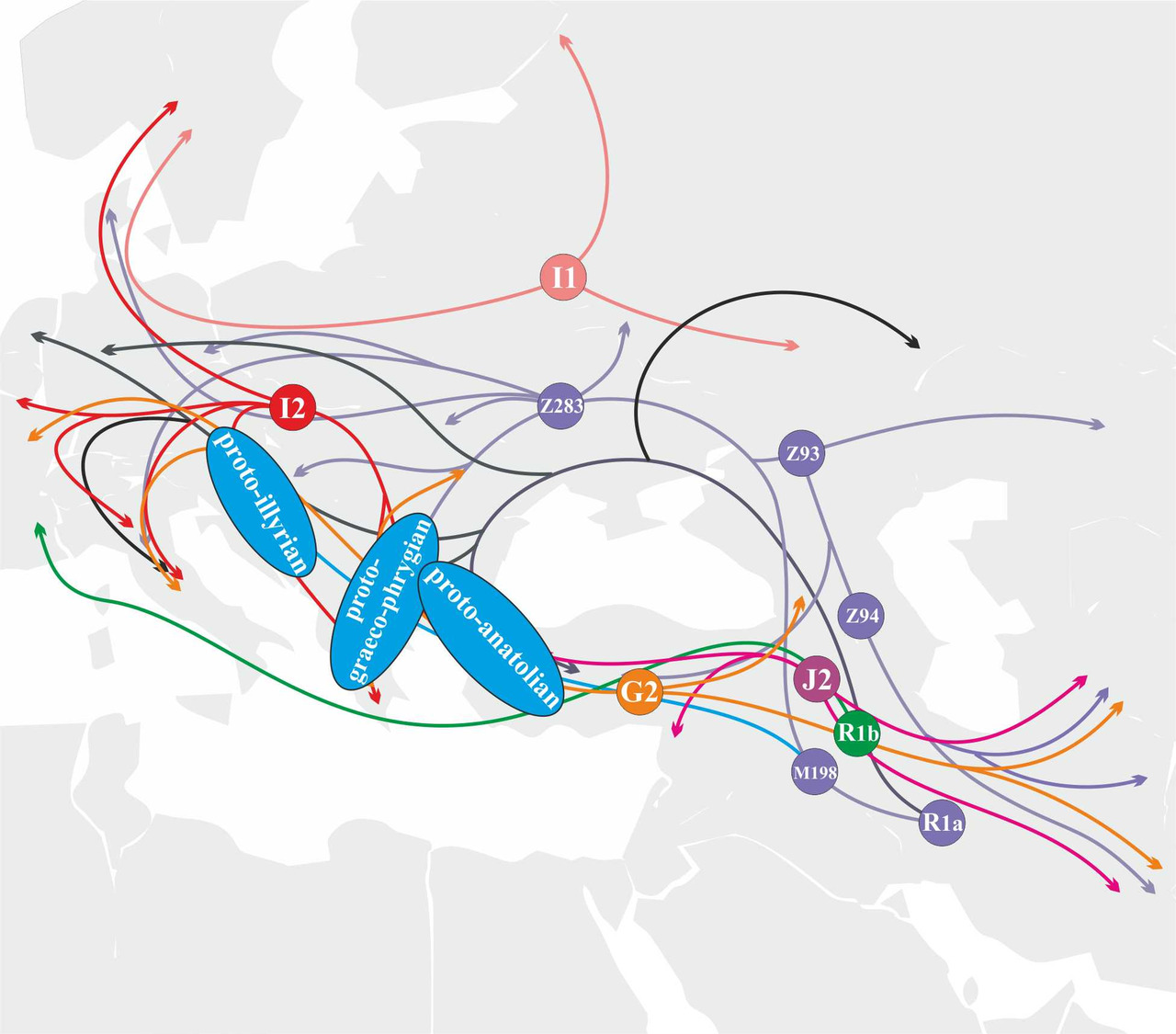
Носители y-гаплогруппы G2 относятся к первой волне неолитических фермеров, пришедших в Европу из Анатолии. Носители y-гаплогруппы G2 принимали активное участие в генезисе ветвей индоевропейских языков, возникших на территории Южной и Центральной Европы, Передней Азии и Иранского нагорья (рис.33).
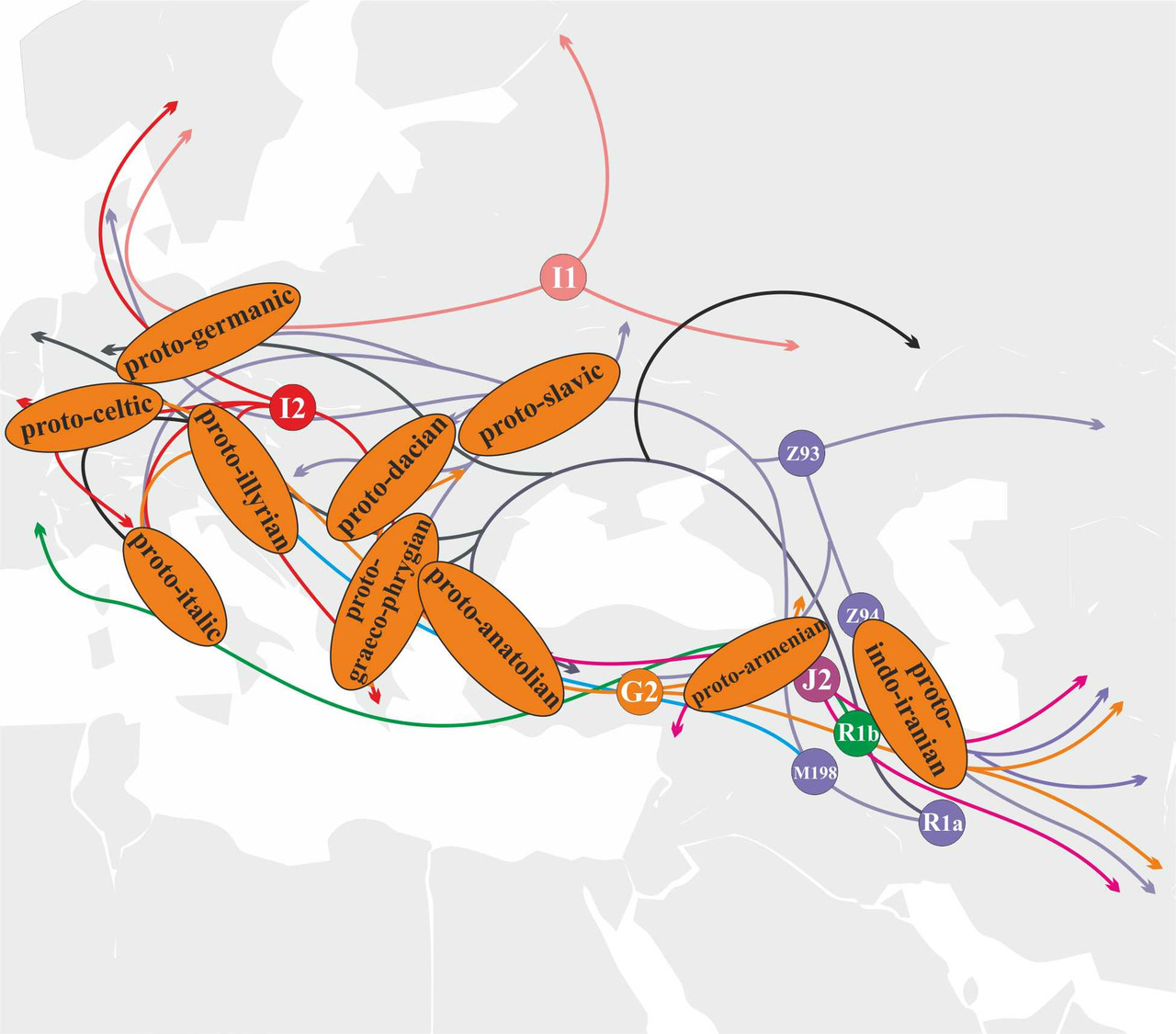
Носители y-гаплогруппы I2 относятся к популяциям, принимавшим активное участие в генезисе ветвей индоевропейских языков, возникших на территории Южной и Центральной Европы (рис.34).
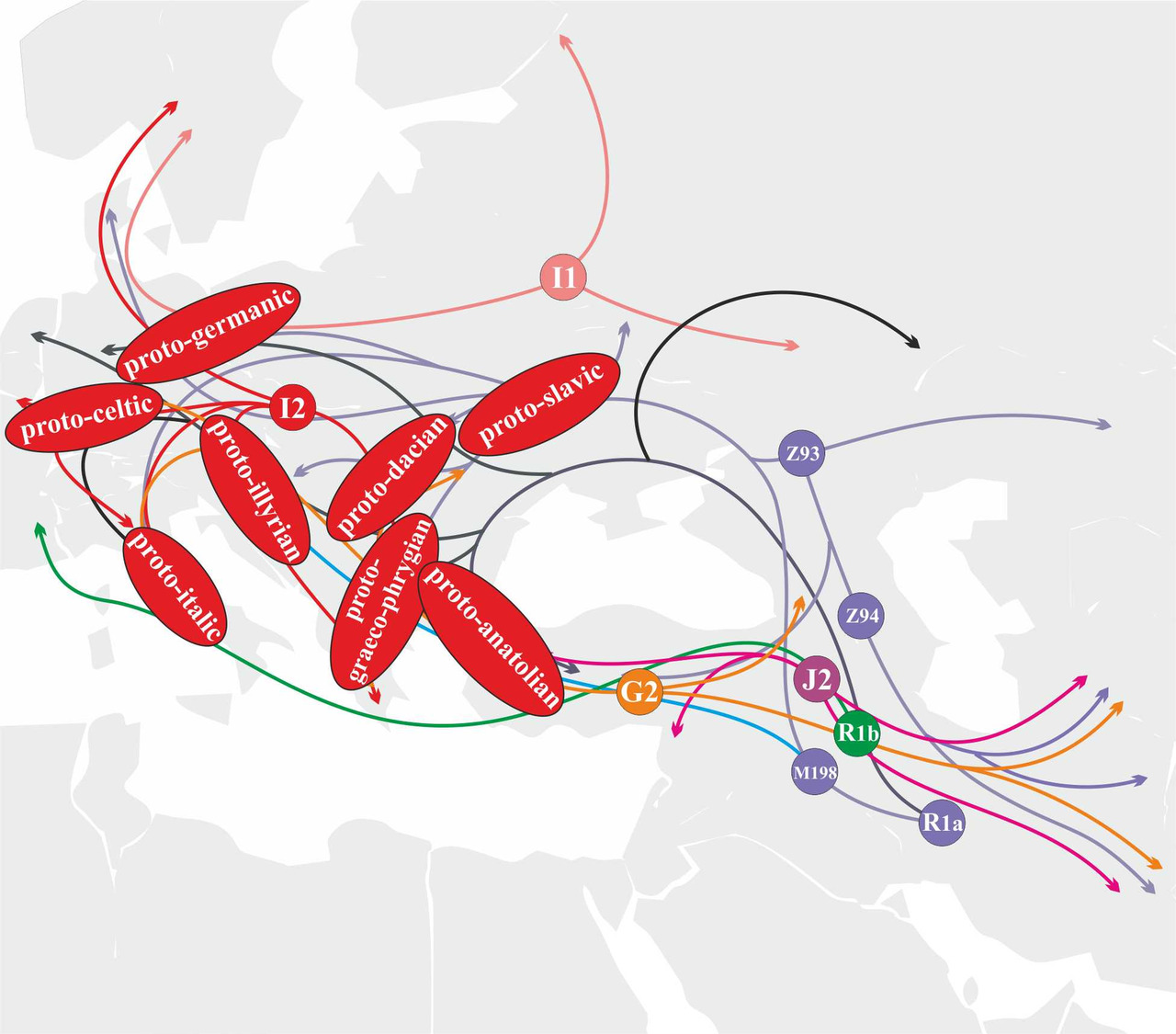
Носители y-гаплогруппы I1, как и носители y-гаплогруппы I2, относятся к старым жителям Европы (кроманьонцам). В результате миграции неолитических фермеров на территорию Европы, а также миграций носителей y-гаплогруппы I2 из Центральной Европы на территорию Балканского полуострова, носители y-гаплогруппы I1 были вынуждены мигрировать в Восточную Европу (днепро-донецкая культурная общность). Носители y-гаплогруппы I1 принимали активное участие в генезисе протогерманского языка (рис.35).
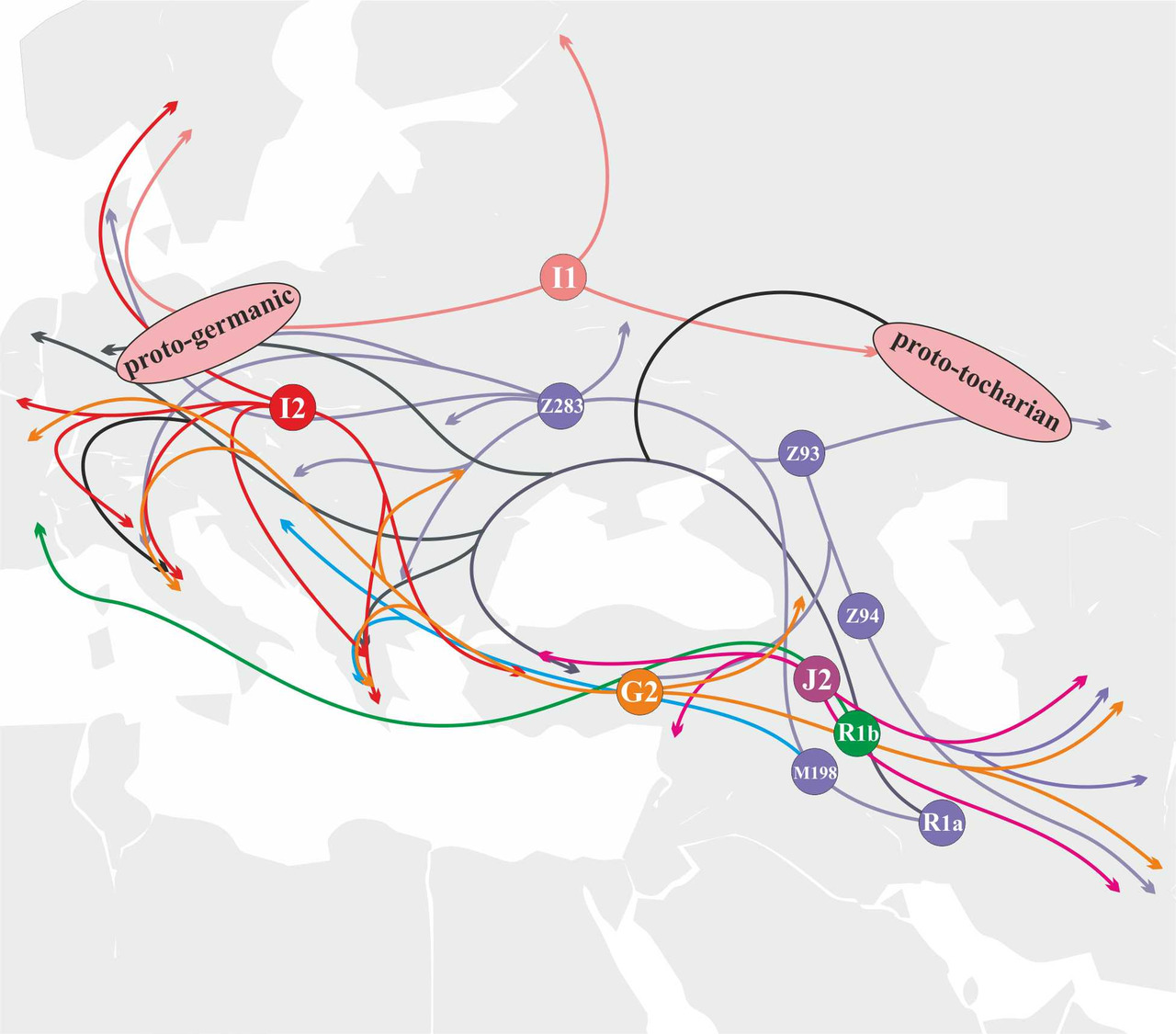
Носители y-гаплогруппы J2 расселялись с территории, которая географически может быть локализована между озерами Ван и Урмия. Носители этой y-гаплогруппы принимали активное участие в генезисе протоиндоиранского и протоармянского языков (рис. 36).
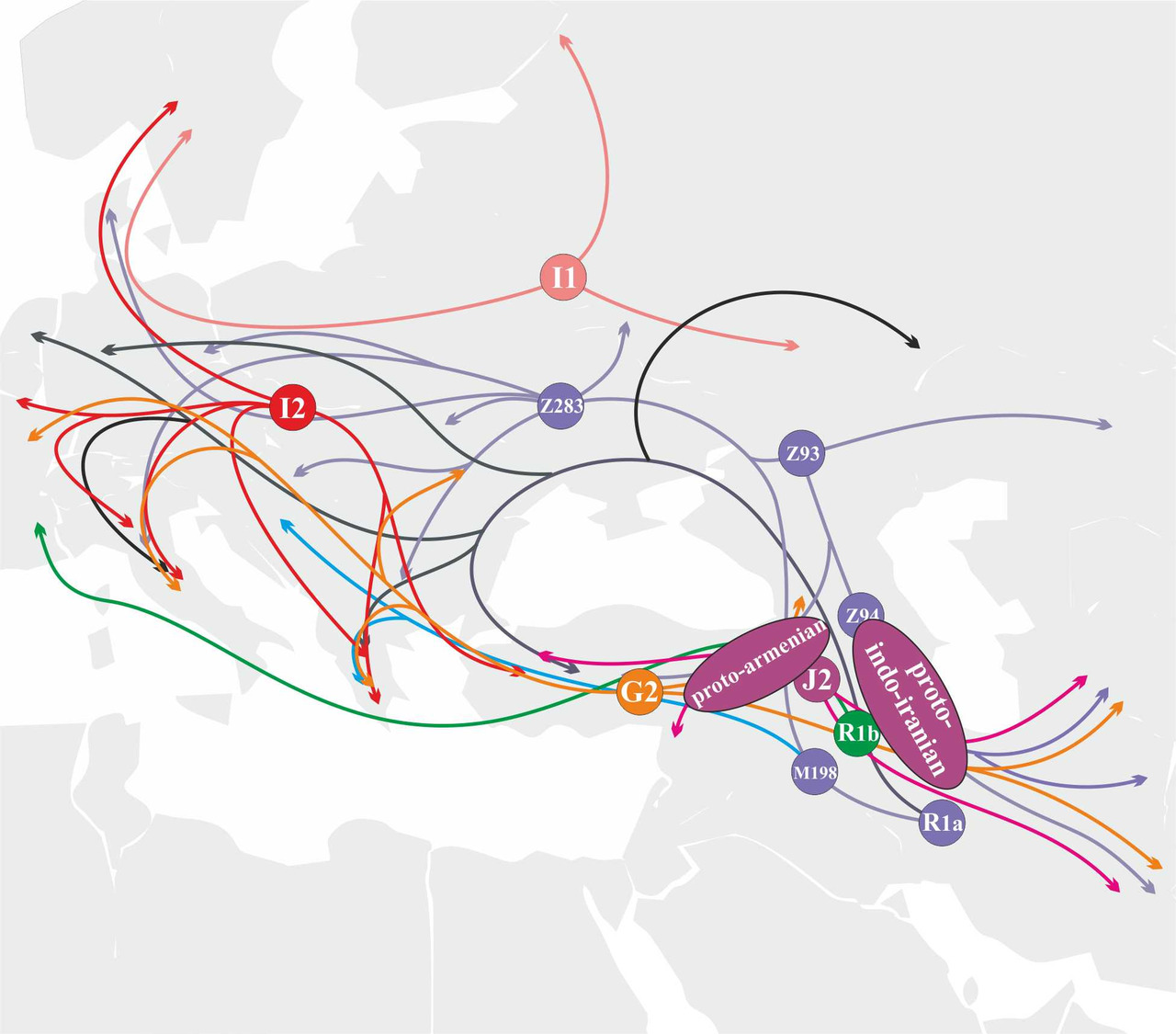
В отличие от носителей y-гаплогруппы R1a, попытки экспансии носителей y-гаплогруппы R1b на территорию Восточной и Центральной Европы в период формирования индоевропейских языков не были столь успешными. Соответственно, участие популяций носителей y-гаплогруппы R1b в генезисе индоевропейских языков является не столь существенным. На сегодняшний день есть основания считать, что носители y-гаплогруппы R1b принимали заметное участие в генезисе протоармянского и протоиталийских языков (рис.37).
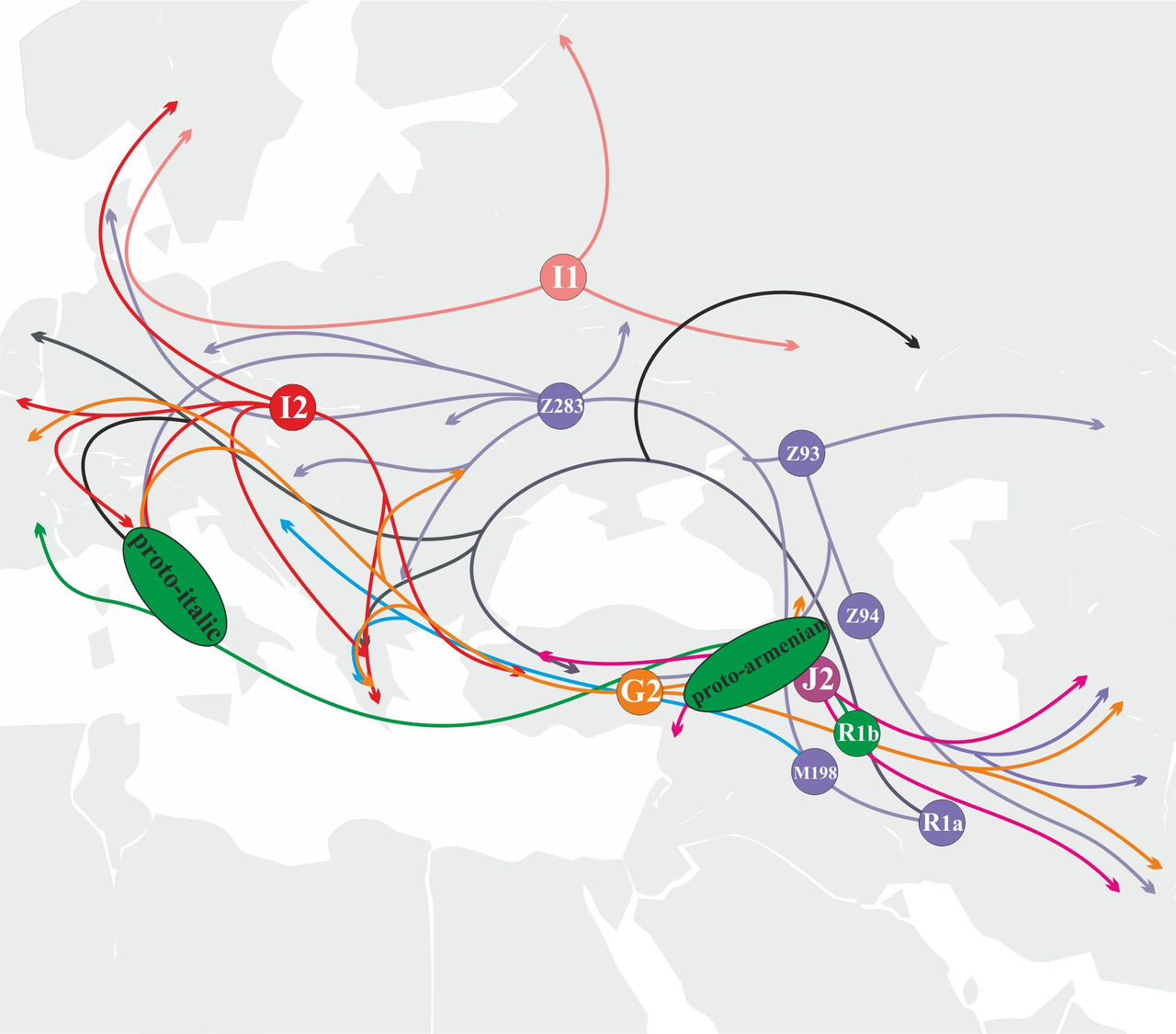
Как можно увидеть из приведенных выше рисунков, основной вклад в генезис индоевропейских языков внесли популяции носителей y-гаплогрупп I2, G2 и R1a. Эти популяции находились в самом центре миграционных процессов. Соответственно, при создании модели генезиса индоевропейских языков на территории Европы можно ограничиться только этими тремя y-гаплогруппами.
Глава III. Моделирование процессов развития языка в популяциях первобытных людей
3.1. Трансформация лингвистических множеств в период формирование макроветвей и изолированного существования популяций
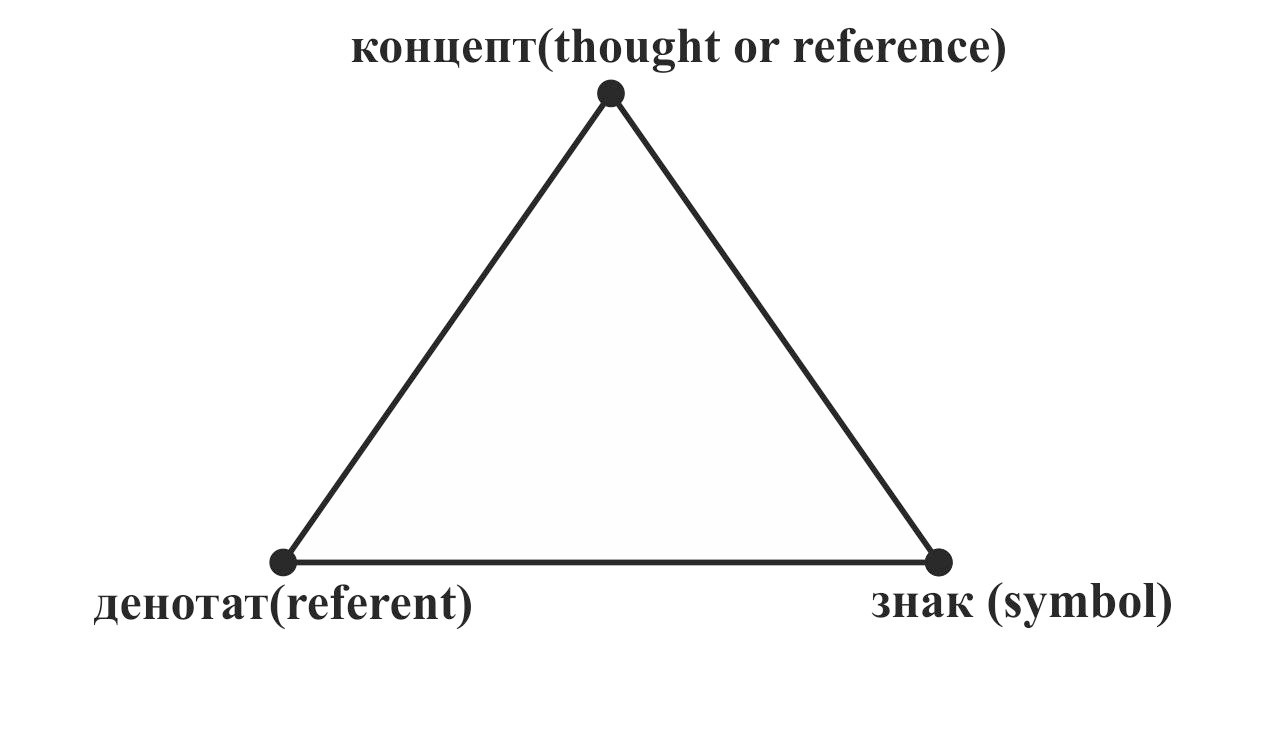
Одной из основных структурных единиц языка является слово. С точки зрения формальной логики слово может быть представлено как величина (структура), характеризуемая тремя основными параметрами: денотат — данный в ощущениях объект реальной действительности или явление психического мира, концепт — возникающий в сознании людей мысленный образ (психологическое представление) о предмете или явлении и знак — акустический образ. Связь между этими тремя параметрами можно продемонстрировать с помощью семантического треугольника Огдена-Ричардса [6] (рис.38).
Лексикон любого языка мы можем представить в виде множества L, состоящего из n-го количества элементов (слов):
L = {slovo1, slovo2, slovo3…, slovon-1, slovon},
где L — множество, включающее в качестве элементов весь лексикон определенного языка;
slovoi — элемент множества L, являющийся величиной, характеризуемой через параметры денотат, концепт, знак (акустический образ);
n — общее количество слов в конкретном лексиконе.
При создании модели трансформации языков, будем считать, что лексиконы всех языков (живых и мертвых) являются результатом трансформации лексикона популяции АСЧ (множество L (CT)), вышедшей с территории Африки ~70 000 лет назад. После возникновения первичных сдвигов согласных звуков в каждой макроветви должно было появиться новое множество, состоящее из слов, акустические образы которых включают согласные звуки, возникшие после первичного сдвига согласных звуков. В этом случае результат трансформации исходного множества L (CT) до уровня формирования макроветвей A-B-C, C-D, G, IJ и K можно представить как сумму нового множества, возникшего в каждой макроветви, и множества L* (CT), являющегося результатом трансформации исходного множества L (CT) (рис.39).
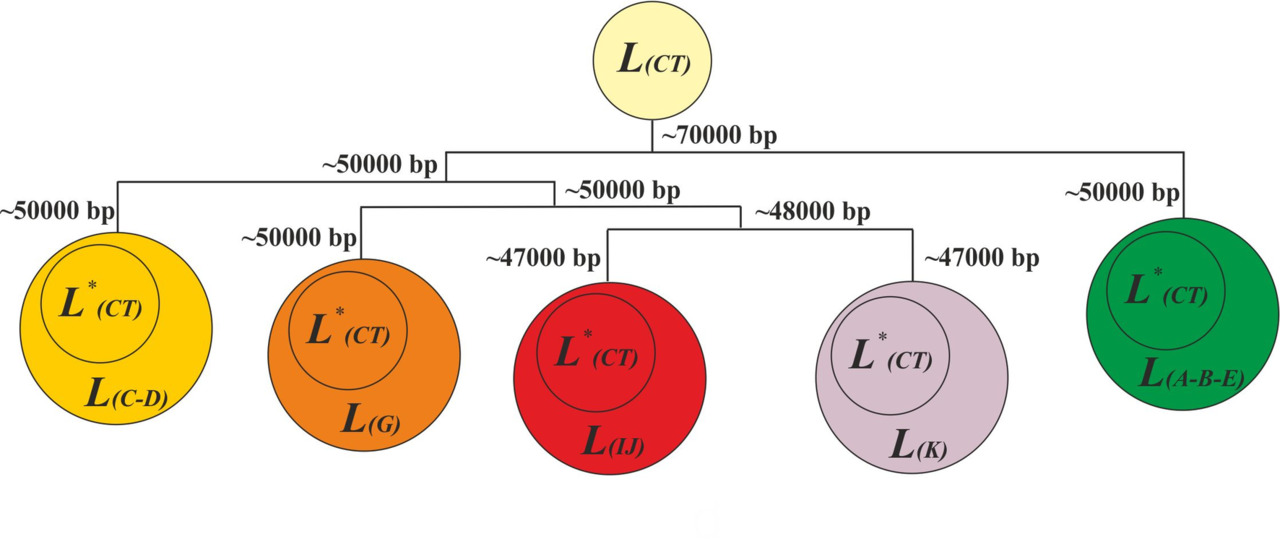
При создании модели трансформации исходного множества L (CT) будем исходить из предположения, что количество слов в лексиконе первобытных людей на уровне y-гаплогруппы CT было незначительным (несколько десятков) и после первичных сдвигов согласных звуков акустические образы данных слов были практически полностью заменены на новые акустические образы. Другими словами, будем считать, что в результате трансформации до уровня формирования макроветвей акустические образы слов исходного множества L (CT) были практически полностью заменены на новые акустические образы. Соответственно, схема трансформации исходного множества L (CT) до уровня формирования макроветвей может быть представлена следующим образом (рис.40).
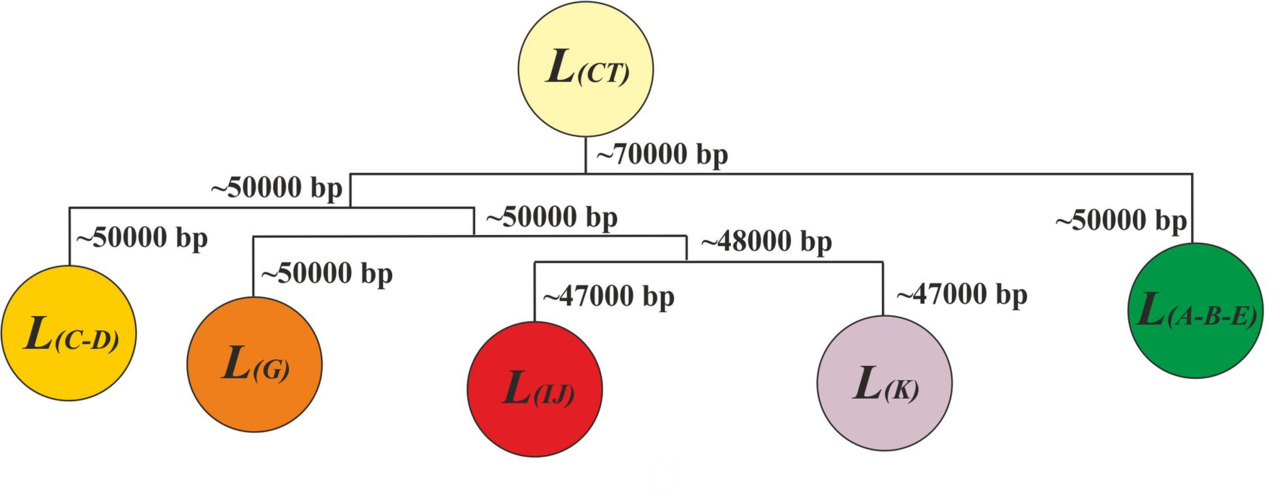
Таким образом, результат трансформации исходного множества L (CT) в период с ~ 45 000 по 8 000 гг. до н.э. (период изолированного существования макроветвей) можно представить в виде:
L (СT) -> L (A-B-E) + L (С-В) + L (G) + L (IJ) + L (K)
В течение всего периода изолированного существования макроветвей шел процесс образованию внутри каждой макроветви множества подветвей. В результате исходное множество слов внутри каждой отдельной макроветви трансформировалось в новые множества, в которых сохранился первичный сдвиг согласных звуков специфичный для исходной макроветви.
Процесс трансформации исходного лингвистического множества в упрощенном виде можно представить как процесс создания новых слов и изменение параметров уже существующих слов. Попробуем смоделировать основные процессы, связанные с созданием новых слов и увеличением лексикона, в период изолированного существования первобытных популяций. Предположим, что в начальный момент времени t1 мы имеем лексикон, состоящий из множества элементов L (t1). Через некоторый промежуток времени в момент времени t2 первоначальное множество L (t1) трансформируется в множество L (t2) (рис.41).
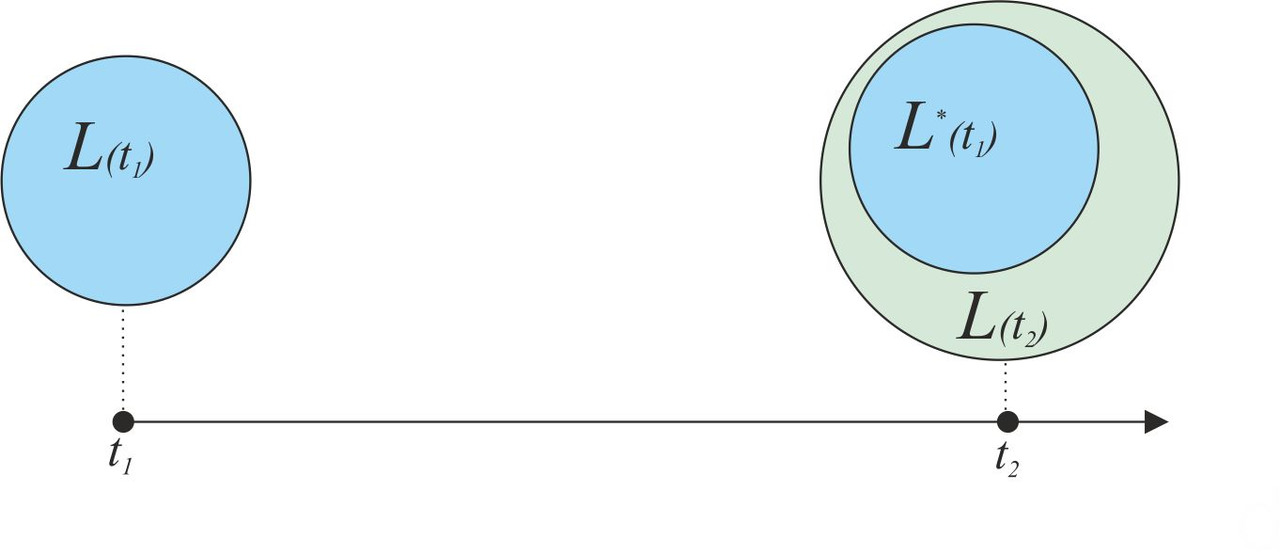
Множество L (t2) будет состоять из множества элементов L* (t1), являющегося результатом трансформации исходного множества L (t1), и множества новых элементов ΔL, возникшего в соответствующий временной интервал Δt= t2- t1. Проанализируем возможные изменения элементов множества L (t1) при трансформации в множество L* (t1). Предположим, что множество L (t1) включало в себя в качестве элемента slovoi, которое трансформировалось за период времени Δt в соответствующий элемент slovo*i. При этом под трансформацией элемента slovoi мы подразумеваем изменение его исходных параметров (денотат, концепт, акустический образ) (рис. 42).
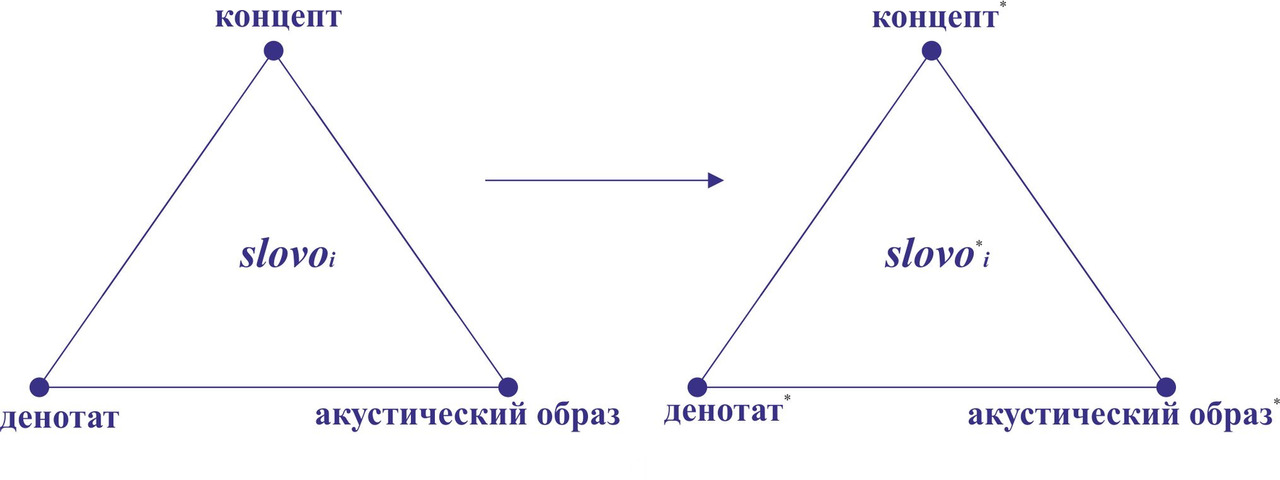
Значения параметров денотат и концепт для большинства предметов и явлений окружающего мира с течением времени будут оставаться неизменными, т.е. в подавляющем числе случаев мы можем поставить знак равенства между значениями параметров денотат и денотат* и концепт и концепт*. В тоже время вероятность изменения акустического образа элемента slovoi при значительном временном промежутке Δt будет достаточно высокой. Таким образом, из трех параметров элемента slovoi наименее стабильным будет акустический образ, т.е. вероятность изменения акустического образа слова будет всегда выше вероятности изменения его денотата и концепта.
Процесс создания новых слов (множество ΔL) является отражением результата активного познания человеком предметов и явлений окружающего мира. Новые знания должны быть зафиксированы в новых словах. Учитывая, что новые знания всегда лежат на фундаменте знаний, накопленных и зафиксированных в словах предыдущими поколениями, вполне логично предположить, что большая часть акустических образов новых слов будет являться производными от акустических образов базового лексикона (рис.43).
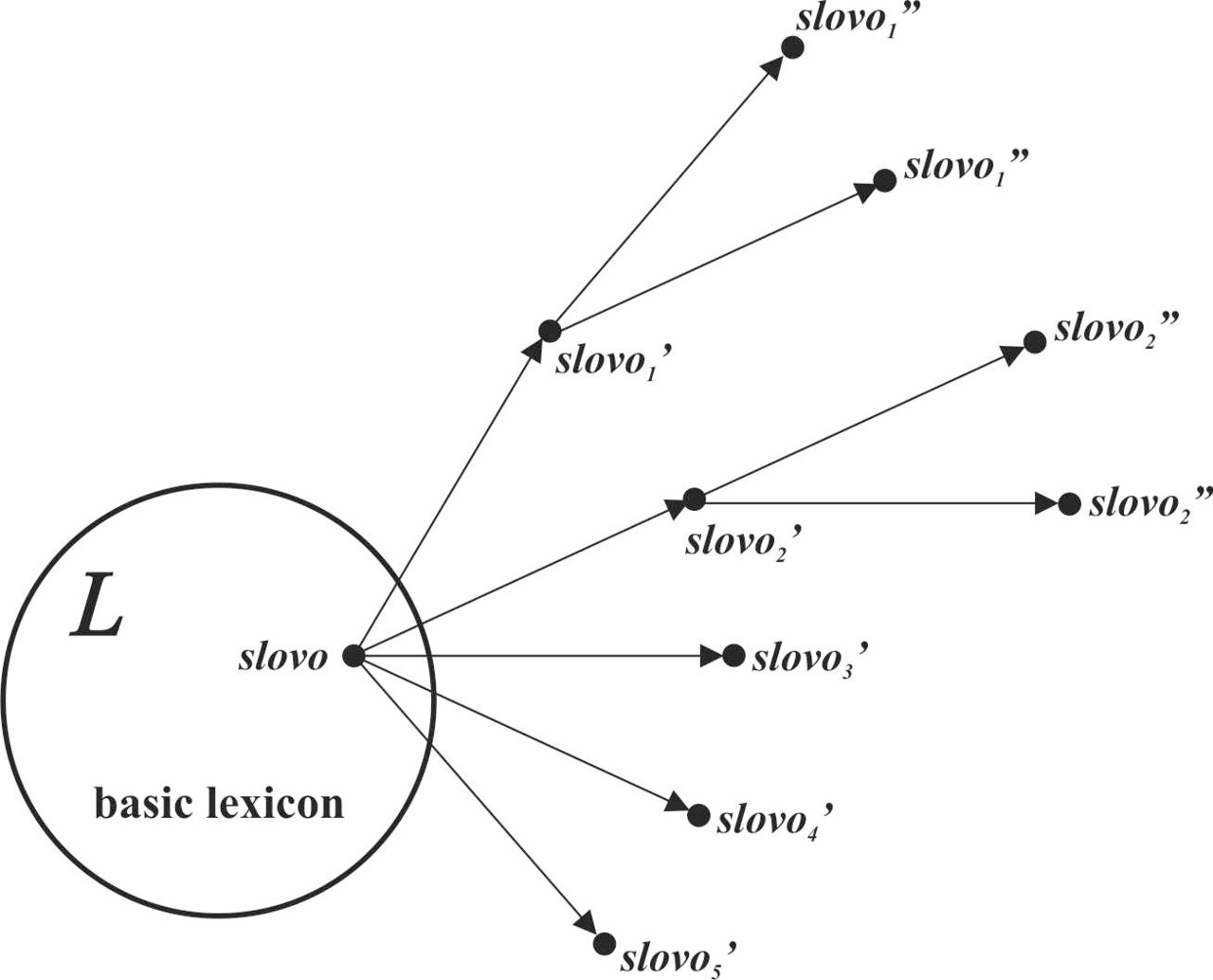
В силу доминирования в базовом лексиконе акустических образов, состоящих из специфичных для соответствующей макроветви согласных звуков, в процессе увеличения лексикона будет воспроизводиться специфичный для базового лексикона сдвига согласных звуков. Таким образом, основные изменения при трансформации лингвистических множеств в период изолированного существования макроветвей можно продемонстрировать следующей схемой (рис.44).
3.2. Трансформация лингвистических множеств в период смешения популяций
При смешении двух популяций, говорящих на языках, относящихся к разным макроветвям, возникнет гибридный язык, который будет частично включать лексикон каждой из исходных популяций, а также новые слова, возникшие уже после гибридизации. Проанализируем теоретически возможные процессы, связанные со смешением двух популяций, говорящих на языках, принадлежащих к двум разных макроветвям. Предположим, что происходит гибридизация носителей y-гаплогруппы I1 (множество L (I1), макроветвь IJ) и y-гаплогруппы R1a (множество L (R1a), макроветвь K) (рис.45)
В процессе гибридизации популяций носителей y-гаплогрупп R1a и I1 должны были произойти следующие основные события:
а) появление гибридных множества L* (I1) и L* (R1a) (рис.46).
Каждое из гибридных множеств L* (I1) и L* (R1a) должно было состоять, как минимум, из четырех основных подмножеств:
ΔL (I1) + ΔL (R1a) +LH (I1+R1a) + LH (R1a+I1)
где ΔL (I1) — часть множества L (I1), вошедшая в гибридное множество;
ΔL (R1a) — часть множества L (R1a), вошедшая в гибридное множество;
LH (I1+R1a) — множество новых слов с акустическими образами, созданными популяцией носителей y-гаплогруппы I1 с использованием согласных звуков специфичных для макроветви K;
LH (R1a+I1) — множество новых слов с акустическими образами, созданными популяцией носителей y-гаплогруппы R1a с использованием согласных звуков специфичных для макроветви IJ;
б) обмен согласными звуками между популяциями (рис.47);
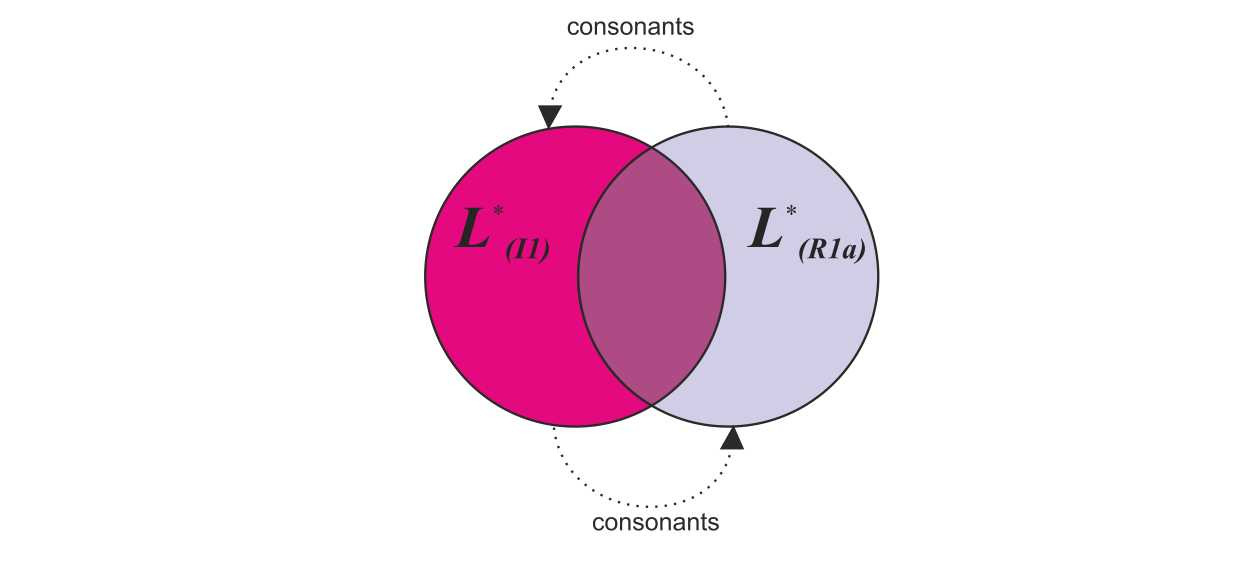
При заимствовании согласных звуков будут происходить их искажения (вторичные сдвиги согласных звуков).
в) Сохранение популяциями первичных сдвигов согласных звуков в процессе гибридизации (рис.48).
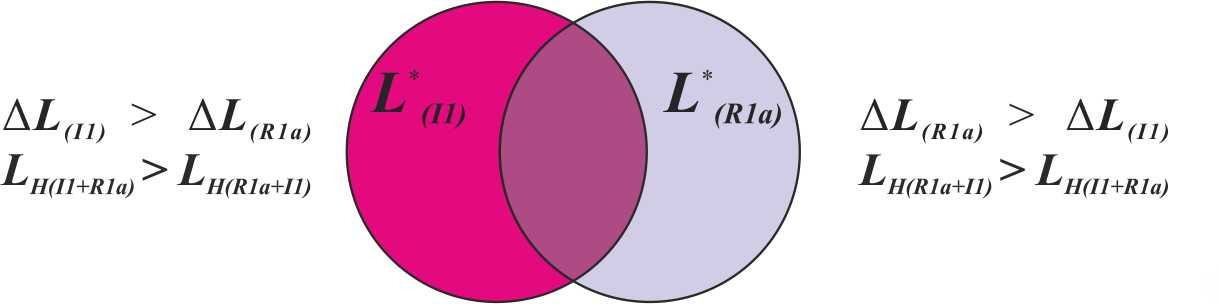
В популяции, состоящей преимущественно из носителей у-гаплогруппы R1a, будет наблюдаться первичный сдвиг согласных звуков специфичный для макроветви K. При этом для гибридного множества L* (R1a) будут справедливыми следующие неравенства: ΔL (R1a)> ΔL (I1), LH (R1a+I1)> LH (I1+R1a). В тоже время в популяции, состоящей преимущественно из носителей у-гаплогруппы I1, будет наблюдаться первичный сдвиг согласных звуков, специфичный для макроветви IJ. При этом для гибридного множества L* (I1) будут справедливыми следующие неравенства: ΔL (I1)> ΔL (R1a), LH (I1+R1a)> LH (R1a+I1).
г) возникновение вторичных сдвигов согласных звуков. Вторичные сдвиги согласных звуков будут зависеть от популяций, принимающие участие в гибридизации.
Таким образом, основные изменения при трансформации лингвистических множеств в период смешения популяций можно продемонстрировать следующей схемой (рис.49)
Глава IV. первичные сдвиги согласных звуков для макроветвей IJ, G и K
4.1. определение первичных сдвигов согласных звуков для макроветвей IJ, G и K
Сама по себе задача определения первичных сдвигов согласных звуков не является сколько-нибудь сложной и может быть выполнена путем анализа акустических образов базового лексикона индоевропейских языков. Первичные сдвиги согласных звуков в популяциях носителей индоевропейских языков должны коррелироваться со структурой y-гаплогрупп в соответствующих популяциях. Однако проведение анализа сдвигов согласных звуков по современным популяциям нельзя считать полностью корректным. Структура y-гаплогрупп популяции может существенно меняться с течением времени. Соответственно, существует риск, что после окончания процесса гибридизации и формирования определенной языковой общности структура y-гаплогрупп популяции претерпела существенные изменения. Учитывая данное обстоятельство, при определении первичных сдвигов согласных звуков необходимо ограничиться источниками акустических образов, которые можно условно считать возникшими до смешения популяций. К таким источникам можно отнести названия водных объектов (гидронимы). Выбор гидронимов в качестве источников акустических образов для определения первичных сдвигов согласных звуков является идеальным по следующим причинам:
а) Для определения первичных сдвигов согласных звуков подходят только акустические образы, которые должны были с наибольшей вероятностью существовать в базовом лексиконе первобытных людей. Учитывая зависимость человека от воды и широкую распространенность воды в природе, будет логичным предположить, что слово для обозначения воды появилось в лексиконах популяций первобытных людей одним из первых. Принадлежность слова к базовому лексикону является гарантией того, что его акустический образ включает согласные звуки специфичные для соответствующей макроветви;
б) Источником названий водных объектов на соответствующей территории, как правило, являлась популяция, пришедшая на данную территорию раньше других. Таким образом, должна существовать корреляция между распространением определенных гидронимов и носителями соответствующих y-гаплогрупп;
в) Названия водных объектов, скорее всего, являлись производными от акустического образа, который использовался соответствующей популяцией для обозначения воды. Соответственно, можно предположить, что названия водных объектов, скорее всего, являются производными от акустического образа, который популяция, проживавшая рядом с этими водными объектами, использовала для обозначения воды. Таким образом, анализ гидронимов, расположенных на территориях проживания носителей соответствующих y-гаплогрупп, дает возможность реконструировать исходные акустические образы, которые использовались этими популяциями для обозначения воды.
При выборе территории, на которой будут анализироваться гидронимы, ограничимся территорией, на которой проживали популяции, принимавшие участие в генезисе индоевропейских языков (носители y-гаплогрупп I1, I2, J2 и G2 и R1) (рис.50).
В качестве материала для анализа возьмем названия крупных рек, протекающих на выбранной территории (рис.51).
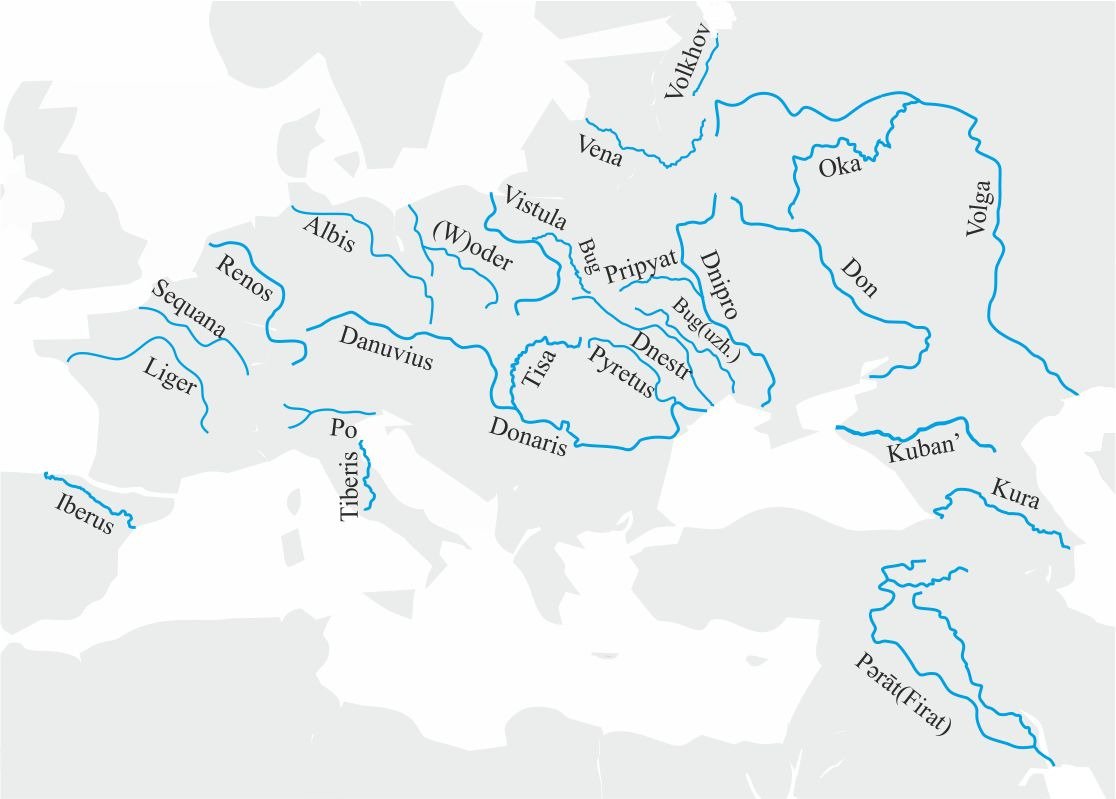
Основным критерием при выборе названий рек будет критерий автохтонности популяции, которая предположительно дала название водному объекту. Соответственно, если какая-либо река имеет несколько названий, оригинальным будет считаться название, которое было использовано для ее обозначения самой ранней популяцией, жившей на территории, по которой протекала река. Например, в случае с рекой Висла таким названием будет Вистула (Vistula), а не Висла, а в случае с рекой Западная Двина таким название будет Вена (Vena). Если название реки, данное автохтонной популяцией, неизвестно, будет использоваться латинское название реки. Для реки Дунай, учитывая ее протяженность, будут использованы два названия: фракийско-дакское — Donaris и латинское — Danuvius. Название реки Oder восстановим до (W) oder, так как выпадение первого согласного звука в названии реки вполне очевидно. Река Тигр учитываться при анализе не будет, так как существует вполне достоверная этимология названия этой реки.
Соответственно, в качестве материала для анализа были выбраны следующие акустические образы названий рек: Po, Iberus, Volkhov, Volga, Pripyat’, Pyretus, Pǝrāt, Vena, Аlbis, Liger, Lige, Renos, Vistula, Bug, (W) oder, Kura, Oka, Kuban’, Donaris, Don, Tibres, Tisa, Sequana, Danuvius, Dnipro и Dnestr. При анализе названий вышеперечисленных рек будем исходить из предположения, что источником названий этих рек могли быть только популяции носителей y-гаплогрупп IJ (макроветвь IJ), G2 (макроветвь G) и R1 (макроветвь К). В этом случае названия выбранных для анализа рек можно рассматривать в качестве результата гибридизации этих трех популяций (рис. 52).

Результат гибридизации этих трех популяций может быть представлен как сумма следующих подмножеств:
Бесплатный фрагмент закончился.
Купите книгу, чтобы продолжить чтение.