
Существует только один Бог — знания
и только один дьявол — невежество.
Сократ из Аттики
Введение. Определение и задачи биологии как науки
Термин «биология» предложил в 1802 г. создатель первой теории эволюции Жан Батист Ламарк. Дословно, биология — это учение о жизни: «биос» — жизнь и «логос» — учение (с греч.). Наука биология изучает биологическую форму движения материи. Это высшая форма движения материи, т.к. она объединяет в себе другие формы — химическую, физическую, механическую (Ф. Энгельс).
Биология относится к естественным наукам и в настоящее время занимает ведущее место в общей системе знаний, поскольку позволяет наиболее полно раскрыть сущность жизни и управлять жизненными процессами.
Главная задача биологии на современном этапе — это создание рациональных взаимоотношений человека и природы, без чего дальнейшее существование жизни на Земле бесперспективно.
Жизнь на Земле олицетворяет живое вещество, представленное огромным многообразием организмов различной степени сложности. В связи с этим в биологии сложились научные направления и оформились специальные науки: вирусология, микробиология, зоология, ботаника и др. В свою очередь, они включают определенные разделы других наук: генетики, биохимии, биомеханики и т. д.
Таким образом, современная биология — это система биологических знаний. Особое место в этой системе занимает биология человека.
Человек — биосоциальное существо, представляющее неотъемлемую часть природы. Он подчиняется общим принципам и закономерностям жизнедеятельности. Вместе с тем, человеку, популяциям людей и антропобиоценозам свойственны свои параметры развития и существования, которые наряду с биологическими изучают и социальные науки. Теоретические достижения антропобиологии нашли широкое применение в медицине, что привело к формированию медицинской биологии. В реально обозримом будущем медицинская биология и ее составляющие: молекулярная биология, биология развития, клиническая генетика, паразитология, популяционная генетика человека позволят решить целый ряд, казалось бы, неразрешимых ранее проблем, связанных с профилактикой, диагностикой и лечением многих заболеваний, увеличением продолжительности жизни, оздоровлением среды обитания.
Роль биологии на современном этапе столь велика, что по уровню биологических знаний и развитию биотехнологий в той или иной стране судят об ее научно-техническом потенциале и интеллектуальном состоянии общества.
Научно-техническая революция обогатила биологию широким набором методов и методических комплексов исследований. Все они обязательно указываются в специальных руководствах и учебниках по разделам биологических знаний. Их можно сгруппировать как общебиологические методы и методы по разделам основных биологических наук.
I. Общебиологические методы
Наблюдение. Основано на получении информации о природном процессе, явлении или объекте с последующим анализом и выводами. Наблюдения бывают полевые (на полигонах) или лабораторные (in vivo — протекающие в живом организме, in vitro — вне организма). Частный вид наблюдения — мониторинг: слежение на протяжении определенного времени за объектами или явлениями, с использованием различных регистрирующих средств.
Эксперимент. Искусственное воспроизведение процесса или природного явления с регистрацией полученных данных и их последующим анализом. Частный вид эксперимента — моделирование, метод исследования сложных объектов или процессов путем их упрощенного иммитирования. Иммитация может быть натурной, математической, логической. Метод базируется на теории подобия.
II. Методы по разделам основных биологических наук
Цитологические связаны с исследованиями клетки: микроскопия — световая, электронная, ультрафиолетовая; метод культуры ткани, метод микрохирургических воздействий на клетку, гистохимические методы.
Генетические связаны с изучением наследственности и изменчивости: гибридологический, генеалогический, близнецовый, популяционно-статистический, цитологический, биохимический; комплекс селекционных методов.
Методы изучения эволюции: исторический, палеонтологический, сравнительно-анатомический, эмбриологический.
Экологические методы: мониторинг, математическое моделирование, эксперимент.
Биотехнологические методы — комплекс наиболее перспективных методик, характеризующих появление синтетической биологии, позволяющей создавать новые биологические компоненты, устройства и системы.
Раздел І. Биологические основы жизнедеятельности. Общая характеристика жизни
Природа жизни
Жизнь как биологическая форма движения материи — одна из наиболее сложных проблем Мироздания. Она объективно существует на планете Земля в течение долгого исторического периода. Одним из первых ученых, представивших основы планетарно-космической организации жизни, был отечественный академик В. И. Вернадский.

Согласно разносторонним оценкам, возраст Земли составляет примерно 4,5—5,0 миллиардов лет. Жизнь на Земле длится около 4,0 миллиардов лет. Таким образом, становление нашей планеты и возникновение на ней жизни в космических параметрах времени произошло почти одновременно. Очевидно, что дальнейшая эволюция происходила при их тесном взаимодействии, нося взаимообусловленный характер. Биолог и геохимик В. И. Вернадский глубоко осмыслил это явление. Им была создана новая обширная область знаний, которую теперь называют наукой о Земле. В ней взаимодействуют геология, геохимия и гидрохимия, почвоведение, география и, конечно, биология. Принципиально новый подход состоял в том, что ученый объединил биоту — живое вещество и сферу его обитания — косное вещество в единое целое — биосферу, живую оболочку Земли.
Живое вещество представлено всей совокупностью живых организмов планеты, существующих в данный момент, независимо от систематики. Оно биохимически чрезвычайно активно и связано с неживой природой непрерывными биогенными потоками атомов и молекул при реализации своих основных функций: питание, дыхание, выделение, размножение. Возникнув, живое вещество приобрело и совершенствовало уникальную способность улавливать, аккумулировать и трансформировать космическую энергию Солнца. Тем самым, в ходе эволюции Земли возник мощный фактор, определивший ход последующих глобальных перестроек ее поверхности. Как отметил В. И. Вернадский, весь лик Земли на современном историческом этапе — ее ландшафты, газовый состав атмосферы, химизм океанов — результат работы живого вещества. Оно придало планете Земля уникальность не только в масштабах Солнечной системы, но, возможно, и Галактики. Идею космической организации жизни во всех ее разнообразных проявлениях, включая человечество, отражают следующие слова выдающегося естествоиспытателя: «Отдельные частные явления соединяются вместе как части одного целого, и в конце концов получается одна картина Вселенной, Космоса, в которую входят и движение небесных светил и строение мельчайших организмов, превращения человеческих обществ, исторические явления, логические законы мышления или бесконечные законы формы и числа» (В. И. Вернадский «Труды по всеобщей истории науки», М., 1988, С. 51—52).
Многообразие проявлений жизни, ее зависимость от положения в Солнечной системе и параметров солнечной активности затрудняет исчерпывающую формулировку. В максимальном приближении жизнь — это глобальная планетарная самоуправляемая энергетически и информационно открытая материальная система, представленная большим разнообразием форм единого в физико-химическом отношении живого вещества.
Главные свойства жизни
В состав живых организмов на атомном уровне входят те же химически элементы, что и в состав неживой материи. Однако, на молекулярном уровне возникают различия, выделяющие живое из неживого.
Живые организмы имеют свойственные только им системы химических связей и взаимодействий между молекулами: ковалентные, ионные, водородные связи, гидрофобные взаимодействия. Биополимеры живых организмов способны образовывать интерполимерные комплексы как между отдельными частями молекул, так и между разными молекулами. Возможность образования этих комплексов, их последующие превращения, а также разрушение обеспечивает важнейшее свойство живой системы — обмен веществ. Его содержание составляют синхронизированные процессы ассимиляции (процессы синтеза, анаболизм) и диссимиляции (процессы распада, катаболизм). В ходе ассимиляции создаются или обновляются различные морфологические структуры, процесс идет с поглощением энергии и называется пластический обмен. При диссимиляции происходит расщепление сложных химических соединений на относительно простые, сопровождающееся выделением энергии — энергетический обмен. Пластический и энергетический обмены тесно взаимосвязаны, составляя единый метаболический цикл. Местом приложения его является клетка.
Живая клетка — это упорядоченная система, в которой природа объединила форму и процесс, структуру и функцию.
Клеткам живых организмов свойственно получать извне, пропускать и частично выделять различные химические соединения. Следовательно, клетки — это открытые системы. Их работа построена по принципу саморегуляции, которая генетически запрограммирована. Хранение генетической информации и ее последующая реализация в длинном ряду поколений осуществляется системой клеточных нуклеиновых кислот. В целом это обеспечивает фундаментальное свойство жизни — историческую непрерывность биологических процессов.
Одним из обязательных свойств жизни является способность к репродукции (размножение). В ходе размножения организмы дают потомство, то есть производят себе подобных. Тем самым, обеспечивается преемственность между родителями и потомками, реализуется механизм наследственности.
Однако, сходство родителей и потомков не бывает абсолютно полным. В этом состоит механизм изменчивости и связанные с ним эволюционные перестройки.
Жизненным отражением космических процессов является эволюционно выработанная биологическая ритмичность — универсальная особенность жизни.
Биоритмы — это количественные и связанные с ними качественные смены биологических процессов, которые проходят на разных уровнях организации. Возникновение их обусловлено планетарными взаимодействиями, вращением Земли вокруг своей оси и вокруг Солнца. Различают экзогенные и эндогенные биоритмы, соответственно вызываемые внешними и внутренними ритмогенными факторами. Самым распространенным является циркадианный (циркадный) суточный биоритм, вытекающий из фотопериода — смены дня и ночи. Растительный и животный мир реагирует на фотопериод фотопериодизмом, сложным комплексом изменений жизнедеятельности. Фотопериодизм служит существенным компонентом таких элементов высшей нервной деятельности как инстинкты. Хронобиология — наука, изучающая закономерности периодически повторяющихся биологических процессов в живом мире.
Важным проявлением жизни служит раздражимость — способность живых организмов реагировать на определенные воздействия окружающей среды. Характер раздражителей, а, следовательно, и ответные адекватные реакции организмов на них очень разнообразны. Они имеют свои особенности у представителей животного и растительного мира.
Наиболее распространенной формой проявления раздражимости служат разнообразные движения — активные и пассивные. Движение — одно из наиболее ярких свойств жизни. В мире животных движения выражены в виде таксисов. Это определенное положительное или отрицательное перемещение относительно раздражителя (фототаксис, термотаксис, хемотаксис). Для растений характерны тропизмы, настии, нутации. В целом движения отражают разные пути эволюционных перестроек и адаптации к среде обитания.
Свойствами живых систем является также дискретность и целостность, энергозависимость, способность трансформировать полученную энергию.
Жизнь многолика. Все ее свойства объединяет единый процесс развития, который охватывает неживую природу, живое вещество и человеческое общество.
Уровни организации жизни
Живая природа представляет сложно организованную систему частей, объединенных общей стратегией жизни. В связи с этим в науке сложились представления об уровнях организации живой материи. Уровень организации определяется по двум принципам — временному и территориальному. Это связано с тем, что разнообразные биологические процессы требуют специфических условий и потому совершаются в определенных границах, выразительно отличаясь по скорости течения. При объединении территориального и временного параметра формируется тот или иной уровень организации в виде сравнительно однородного биологического комплекса. Он характеризуется двумя основными показателями: элементарной структурной единицей и элементарным биологическим явлением. В настоящее время принято выделять следующие уровни:
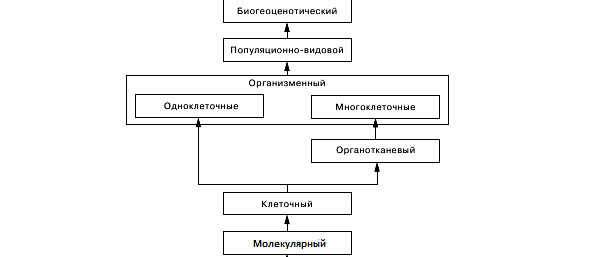
Молекулярно-генетический уровень.
Элементарные структуры — коды наследственной информации, то есть последовательности триплетов нуклеотидов молекулы ДНК. Элементарные явления — воспроизведение этих кодов по принципу матричного синтеза или конвариантная редупликация (удвоение) молекулы ДНК. Механизм редупликации обусловливает тиражирование генов. Это позволяет передавать генетическую информацию в ряде поколений клеток и обеспечивает механизмы наследственности. Имеющие место случаи ошибок синтеза изменяют кодоны, что сразу же воспроизводится в молекулах копиях. Редупликация становится конвариантной, то есть происходящей с изменениями (явления генных мутаций).
Перенос информации в оформленную структуру, белковую молекулу, обеспечивается набором специализированных внутриклеточных образований — органелл в процессе синтеза белка. Экологические проблемы уровня: рост мутагенных воздействий, увеличение доли мутаций в генофондах.
Клеточный уровень. Элементарные структуры — клетки. Элементарные явления — жизненные циклы клеток. Клетка превращает поступающие в организм вещества и энергию в форму, пригодную для использования организмом, тем самым, обеспечивает процессы жизнедеятельности. Каждая клетка относительно автономна и представляет самостоятельно функционирующую единицу. В составе целого организма клетки объединяются в ткани и органные системы. Между ними налажена система физиолого-биохимических и структурно-функциональных связей, характерная для тканей данного организма. Экологические проблемы уровня: рост клеточной патологии в связи с загрязнением среды, нарушение воспроизводства клеток.
Организменный уровень. Элементарные структуры — организмы и составляющие их системы органов. Элементарные явления — комплекс физиологических процессов, обеспечивающих жизнедеятельность. На данном уровне осуществляется механизм адаптации и складывается определенное поведение живых существ в конкретных условиях среды обитания. Наследственная информация, заключенная в генотипе, реализуется теми или иными фенотипическими проявлениями. Управляющая система — генотип. Экологические проблемы уровня: снижение адаптационных возможностей организмов, развитие пограничных состояний у человека (состояние между здоровьем и болезнью).
Популяционно-видовой уровень. Элементарные структуры — популяции. Элементарные явления — видообразование на основе естественного отбора. Популяция — основная эволюционная единица. Важнейший эволюционно-генетический показатель популяции — её генофонд. Это управляющая подсистема уровня. Генофонд определяет эволюционные перспективы и экологическую пластичность популяций. Имеется ряд факторов, вызывающих изменение генофонда популяций: мутации, генетическая комбинаторика, популяционные волны, изоляция. Реализация изменений происходит путем естественного отбора. Экологические проблемы уровня: ухудшение экологических показателей популяций (численность, плотность, возрастной состав и т.д.).
Биосферно-биогеоценотический уровень. Элементарные структуры — биогеоценозы. Элементарные явления — динамическая взаимосвязь биогеоценозов в масштабах биосферы. Управляющая подсистема уровня — генопласт (термин ввел академик М. А. Голубец). Это совокупность генофондов и генотипов, адаптированных друг к другу популяций в окружающей их среде. Весь комплекс биогеоценозов образует живую оболочку Земли — биосферу. Между биогеоценозами происходит не только материально-энергетический обмен, но и постоянная конкурентная борьба, что придает биосфере в целом большую динамичность. Вся биогеохимическая работа биосферы обеспечивается ее биогеоценозным комплексом. Экологические проблемы уровня: увеличение числа антропоценозов и их глобальное распространение, загрязнение сред, изменение озонового экрана Земли.
На любом уровне живой системы происходит поступательное развитие с обменом веществом, энергией, информацией.
Рассмотренные биологические уровни живой природы взаимосвязаны по принципу соподчинения или биологической иерархии. Система низшего уровня обязательно включается в уровень вышестоящего порядка. Следовательно, идея биологических уровней с одной стороны подразделяет живую природу на отдельные составляющие — дискретные единицы, а с другой стороны объясняет ее целостность как системы взаимосвязанных частей, начиная от органических макромолекул и кончая живой оболочкой Земли биосферой.
Формы жизни
В настоящее время на планете Земля сложились две формы жизни — неклеточная и клеточная.
Неклеточная форма жизни
Ее представляют вирусы, которых выделяют в самостоятельную группу Вира (Vira). Открытие вирусов связано с именем русского ученого Димитрия Ивановского. В 1892 году им доказано, что вирусы — это субмикроскопические частицы, способные проходить через фильтры и вызывать инфекционные заболевания. Однако, только в 1935 году основатель вирусологии американец Уэнделл Стэнли получил в чистом виде «дедушку» всех вирусов — вирус мозаичной болезни табака. Он установил белковую природу вируса, способность его белков принимать кристаллическую форму.
Вирус парадоксален! Пребывая в состоянии гигантской макромолекулы (частицы), он не проявляет признаков жизни. Живые свойства вируса обнаруживаются только при попадании его в клетку. Здесь он способен, как все живое, быстро размножаться и мутировать, используя структурные компоненты клетки хозяина. Если в инертном состоянии вирус сохраняется годами и даже веками, то его жизненная форма может существовать лишь несколько часов.
Таким образом, вирусы заполняют сумеречную зону жизни. Они расположились на полпути между молекулой и клеткой, живым и неживым.
Вирусы — это паразиты клеток. Каждый вид вирусов может заражать только определенный клеточный набор. В связи с этим выделяют: вирусы растений, животных (включая человека), грибов и бактерий. Последние называются бактериофаги.
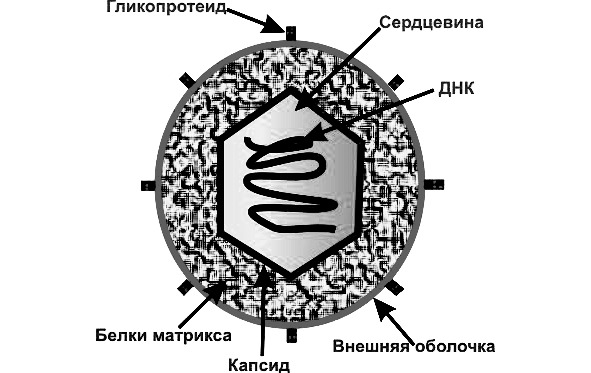
Строение вирусов. Зрелые частицы вирусов — вирионы или вироспоры состоят из белковой оболочки и нуклеокапсида, содержащего генетический материал. Иногда под белковым слоем находится слой липидов, что придает капсиду особую прочность и препятствует проникновению ферментов и биологически активных веществ клетки хозяина к вирусу. Это имеет огромное значение для сохранения его функциональной активности. Под оболочкой содержится нуклеиновая кислота — носитель генетической информации вируса.
По своей генетической организации вирусы подразделяются на две группы. Первая группа вирусов включает только ДНК, которая обычно кольцевидно закручена и имеет одноцепочечную структуру, а не двухцепочечную (двойная спираль).
Вторая группа вирусов содержит РНК. Особенность вирусной РНК состоит в том, что наряду с одноцепочечной структурой она может быть двухцепочечной, то есть приобретать вид двойной спирали (ретровирус, вирус раневых опухолей растений и др.) см. рис.1.
Проникновение вируса в клетки хозяина происходит следующим образом. В месте контакта вируса с клеткой возникает впячивание клеточной мембраны. Оно углубляется внутрь клетки и затем отшнуровывается. В цитоплазме клетки образуется подобие вакуоли, которая вместе с капсидом быстро разрушается и вирусная нуклеиновая кислота оказывается в цитоплазме пораженной клетки.
Дальнейшее взаимодействие может осуществляться двумя способами. Первый состоит в том, что РНК вируса с помощью фермента ревертазы связывается с рибосомой клетки хозяина, в результате чего происходит трансляция генетической информации вируса и в клетке продуцируется патологический вирусный белок. Данный путь характерен для вирусов гриппа и ВИЧ. Второй вид взаимодействия — это способность ДНК вируса встраиваться в ДНК клетки и соответственно передавать свою информацию через геном клетки хозяина. Таким образом реализуют собственные возможности вирусы раковых заболеваний (онковирусы).
Проникнув в клетку хозяина, вирусы быстро подавляют ее системы репарации (восстановления) и могут разрушить клетку. Клетки противодействуют вирусам, вырабатывая антивирусные белки-интерфероны.
Вирус, в отличие от многих клеток, абсолютно лишен собственных средств передвижения. Он передается при непосредственном контакте организмов или через переносчиков, которыми могут быть различные представители животного мира. Несмотря на относительную простоту организации, вирусы прочно удерживают свои позиции в природе. Более того, они усиливают свою агрессивность. На это указывает возросшая частота вирусных заболеваний, а также появление новых вирусов, вызывающих тяжелые заболевания человека: вирус иммунодефицита (СПИД), вирус нетипичной пневмонии.
Клеточная форма жизни
Данную форму жизни представляют клетки двух типов — прокариотические и эукариотические.
Относительно просто устроенные прокариотические, безъядерные, клетки возникли в первичном океане жизни примерно 3,5 миллиарда лет назад. Эукариотические клетки, имеющие ядра, образовались позднее. Мы не имеем неоспоримых доказательств происхождения эукариотических клеток из прокариотических. Здесь возможны пока лишь гипотезы. Важнейшая из них гипотеза клеточного симбиоза. Согласно ей эукариотические клетки вначале эволюционного пути были анаэробными организмами. В дальнейшем установился их стабильный симбиоз с бактериями. Считают, что главная окислительная система клеток эукариот — митохондрии, произошла от особого рода фотосинтезирующих бактерий, утративших способность к фотосинтезу и сохранивших только дыхательную цепь. Возможны и другие комбинации разнообразных жизненных начал. Как бы то ни было, создав биологическую модель в виде клетки, природа в дальнейшем широко использовала ее в ходе эволюции, создавая живые формы различной степени сложности.
Сравнительная характеристика прокариот и эукариот

Раздел II. Клеточный и молекулярно-генетический уровни организации живого. Клетка — элементарная структурно-функциональная единица живого
Природа довольно редко и достаточно долго создает принципиально новые биологические конструкции. Она предпочитает усовершенствовать и комбинировать уже существующее. Поэтому возникновение клетки как живой системы нового типа явилось важным этапом эволюции. Клетка стала основой строения и развития организмов, поскольку обладает универсальным набором свойств, обеспечивающих жизнедеятельность. Назовем важнейшие из них.
Клетка — саморегулирующаяся система. Она имеет четко синхронизированный цикл жизнедеятельности. Известно, что органические соединения реагируют друг с другом медленно, а жизнь не может поддерживаться за счет медленных реакций. Поэтому живая клетка выработала особые ускорители реакций в виде ферментов или энзимов. Производительность их огромна: некоторые ферменты в течение одной минуты способны разложить до 5-ти миллионов молекул субстрата при температуре 0ºС. Ферменты это не универсальные наборы. Каждый из них может произвести только одно направленное действие, в связи с чем и существуют тысячи ферментов. В клетке они включаются в работу синхронно, со строгой последовательностью во времени и пространстве. Регуляцию осуществляет сама клетка.
Клетка — самовоспроизводящаяся система. Благодаря способности к различным видам делений клетки могут повторять себя в длинном ряду поколений и дочерних форм, что делает их с философской точки зрения бессмертными.
Клетка — система, способная к самоопределению. Молодые клетки обычно очень похожи друг на друга. Однако, в ходе развития каждая клетка идет своим путем. Происходит дифференцировка клеток, связанная с тем, что они должны выполнять строго определенные функции в составе многоклеточного организма. Так, в теле человека насчитывается около 250 типов клеток, причем, каждый тип имеет свое назначение.
Клетка — самовосстанавливающаяся система. Наличие молекулярных механизмов репарации молекулы ДНК, а также регенерации различных внутриклеточных органелл позволяют клетке с высокой степенью надежности исправлять различные повреждения. Не вызывает сомнений, что способность к самовосстановлению закладывалась постепенно в ходе эволюции и совершенствовалась в связи с усложнением клеток.
Клетка — энергетически открытая система. Нормальная жизнедеятельность любого организма невозможна без поступления веществ и энергии. Клетка обеспечивает преобразование этих веществ в форму пригодную для использования организмом. Она располагает универсальным механизмом белковое синтеза, продуцируя белки для собственных нужд и экспортируя их другим клеткам. На клеточном уровне обеспечивается не только вещественно-энергетический обмен между организмом и средой, но и происходит использование наследственной информации. Наследственность невозможна без обмена веществ. Все явления жизни взаимообусловлены, а объединяет их клетка.
Клетка — устойчивая система, доказательством служит весь ход эволюции. За последние 500 миллионов лет клетки фактически не изменились. Их функционирование осуществляется с помощью довольно ограниченного набора унифицированных молекулярных, надмолекулярных и морфологических структур.
Перечисленные выше качественные особенности позволяют рассматривать клетку как элементарную и незаменимую биологическую систему. Детальное изучение клеточных структур и их взаимодействий привело в середине прошлого века к формированию представлений о клеточном уровне организации. Опираясь на данный уровень, оказалось возможным ввести понятия и о других структурных уровнях организации живого, как надклеточных, так и субклеточных.
Итак, возникновение клетки сыграло решающую роль для прогресса жизни на нашей планете. Оно обеспечило: возможность передачи наследственной информации в рядах поколения живых организмов; смену гибнущих в процессе жизнедеятельности структур новыми; рост и развитие организмов путем формирования тканей, являющихся опорным материалом для различных систем органов и всего организма в целом.
История открытия клетки
Открытие клетки связывают с именем Роберта Гука (1665 г.), работавшего попечителем экспериментов в Лондонском Королевском научном обществе. Как физика Р. Гука заинтересовали механические свойства пробкового дерева: прочность, упругость, легкость, непотопляемость. Решив исследовать пробку под микроскопом, Р. Гук предварительно существенно модернизировал этот прибор, сконструированный ранее любителем-натуралистом Антони ван Левенгуком. По сути дела, им создан прототип современного светового микроскопа.
Микроскопия тонких срезов пробкового дерева показала, что оно состоит из многочисленных пустых ячеек, которые Р. Гук назвал сеllulа (позднее cell). Так родился термин «клетка» — ячейка. Представления же о клетке, как о структурной единице живой природы, складывались постепенно. Обязательность наличия ядра в этой структуре установил Я. Пуркинье и позднее Р. Броун (1830 г.). Первому автору вводят в заслугу также введение понятия «протоплазма клетки». Механизм клеточного деления открыл лейпцигский продавец книг В. Гофмейстер (1849 г.), сделавший первые зарисовки делящихся клеток. Позднее, В. Ру показал универсальность этого процесса для живого и растительного мира. Сложнее обстояло дело с изучением клеточных мембран. Эти тончайшие структуры животных клеток плохо вырисовывались в обычном световом микроскопе. Тем не менее, принцип их работы удалось правильно предсказать. Заключительный каскад открытий связан с описанием внутриклеточных структур. Разные ученые передовых европейских стран выявили самые миниатюрные образования, клеточные органеллы. В историю цитологии вошли имена Ван Бенедена, Бенда, Гольджи, Де Дюву. Клетка предстала перед глазами в законченном виде. Это позволило оформить теорию клетки, то есть показать принцип ее структурной организации. Теория клетки — частное. От частного оставался один шаг к общему — созданию клеточной теории.
Клеточная теория
Клеточная теория является крупнейшим достижением естествознания XIX столетия. Она утвердила единый принцип организации живой природы на клеточном уровне, связала растительный и животный мир. Это одно из наиболее широких обобщений в биологии. В разработку ее внесли вклад несколько выдающихся ученых. Первостепенная роль принадлежит Теодору Шванну, немецкому физиологу и гистологу. В книге «Микроскопические исследования о соответствии в структуре и росте животных и растений» (1839 г.) он показал, что клетка является главной структурной и функциональной единицей всех животных и растительных организмов. Тем самым, была доказана общность строения и развития животных и растений, провозглашено единство органического мира.
Значительный вклад в разработку клеточной теории внес также ботаник Матиас Шлейден, показавший, что растительные и животные клетки гомологичны, то есть имеют одинаковый принцип организации.
В дальнейшем знаменитый немецкий медик, ученый с непререкаемым авторитетом, Рудольф Вирхов провозгласил: «Оmnis cellula e cellulae», что значит: клетка возникает из клетки путем ее деления. Р. Вирхов первым внедрил клеточную теорию в медицину.
Основные положения клеточной теории
1. Клетка — элементарная структурно-функциональная единица живой материи.
2. Клетки различных организмов сохраняют одинаковый принцип строения.
3. Размножение клеток происходит путем деления исходной материнский клетки.
4. Многоклеточные организмы — это совокупность различных клеток, интегрированных в единую систему живого организма.
Со времени создания клеточной теории наши представления о клетке существенно обогатились, однако суть клеточной теории осталась неизменной.
Химическая организация клетки
Живая клетка — это миниатюрный реактор, в котором не затухают химические процессы. Из 109 элементов периодической системы Д. И. Менделеева клетка включает около 70. Количественные соотношения их варьируют В клетках организма человека значительно преобладает четыре элемента: водород—10%, кислород—60%, углерод — 20%, азот — около 3%. Менее 2% приходится на следующие девять элементов: натрий, кальций, фосфор, железо, сера, калий, хлор, кремний, магний. Все указанные элементы составляют группу макроэлементов. Остальные элементы присутствуют в общем количестве 0,1% массы и относятся к микроэлементам: кобальт, цинк, медь, марганец, хром и другие.
Обратим внимание! Все химические компоненты клетки, вне зависимости от их количества, играют важную роль в жизнедеятельности организма.
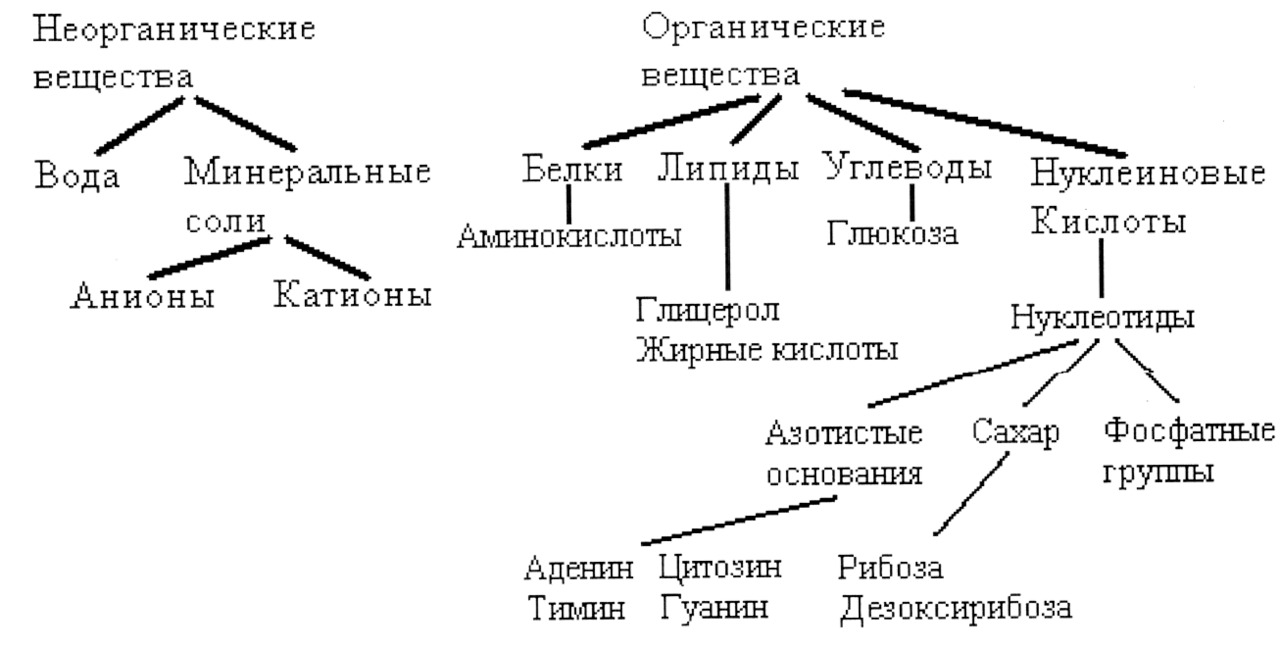
В живой клетке химические элементы находятся либо в виде ионов, либо входят в состав молекул органических или неорганических веществ.
Органические вещества — это углеродсодержащие соединения, присутствующие в живых организмах — отсюда и термин органические. Неорганические вещества характерны для живой и неживой природы и имеют другую химическую организацию.
Неорганические вещества
Вода. Это компонент исключительной важности. Вода составляет 80% всей массы клетки. Всеобщая функция воды в клетках — цементирующая. Она состоит в образовании водородных связей между отдельными частицами и компонентами живой клетки. Молекула воды электрически асимметрична. Это электрический диполь, в котором каждый атом водорода несет положительный заряд, а кислорода –отрицательный. В результате и возникает сильное электростатическое напряжение: Н-связь.
Роль воды в клетке многообразна. Она предопределяет рН среды, от которой во многом зависит работа всех систем органов. Благодаря высокой теплоемкости, вода предохраняет клетку от резких колебаний температуры и способствует теплорегуляции. Это хороший растворитель для многих органических и неорганических веществ, а большинство химических реакций в клетке возможно только между растворенными веществами. Поступление и выведение веществ клеткой осуществляется в водных растворах. Вещества, хорошо растворимые в воде, называются гидрофильными. К ним относятся соли, кислоты, многие спирты. Хорошей растворимостью в воде обладает ряд белков и углеводов. Вещества, плохо растворимые в воде, называют гидрофобными. Среди них на первом месте стоят липиды (жиры). Явление гидрофильности и гидрофобности используется при построении полупроницаемых клеточных мембран. Вода является фактором, определяющим термостабильность клетки.
Минеральные соли. Подавляющая часть неорганических веществ клетки находится в виде солей. Они присутствуют в твердом состоянии или диссоциированы на ионы. Для процессов жизнедеятельности наиболее важны катионы К+, Са2+, Мg2+, анионы НРО4-, Н2РО4-, СL-, НСО3-. Концентрация катионов и анионов в клетке и вокруг нее различна и регулирует поступление воды в клетку. В цитоплазме довольно много К+ и мало Na+. Во внеклеточной среде (морская вода, плазма крови) наоборот много Na+ и мало К+. Эти соотношение быстро выравниваются со смертью клетки.
Органические вещества
Органические вещества в большинстве являются биополимерами. Это белки, углеводы, нуклеиновые кислоты. Биополимеры представляют высокомолекулярные химические соединения, состоящие из относительно однородных мономеров. Мономерами белков служат аминокислоты, углеводов (полисахаридов) — моносахариды, нуклеиновых кислот — нуклеотиды.
Биополимеры подразделяют на информационные: белки, нуклеиновые кислоты и неинформационные — углеводы. Липиды не относят к биополимерам. Указанные четыре класса органических соединений составляют основу живых систем.
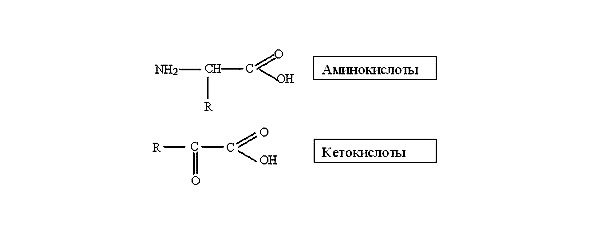
Белки характеризуются высокой молекулярной массой и большим разнообразием. В организме человека насчитывается около 5 млн. типов белковых молекул. Несмотря на столь широкое представительство, белки образованы всего 20 мономерами — аминокислотами.
Двадцать аминокислот в составе белков

Аминокислоты делятся на две группы
1. Незаменимые аминокислоты не могут синтезироваться животными организмами и обязательно поступают с пищей (аргинин, валин, гистидин, изолейцин, лейцин, метионин, фенилаланин, треонин).
2. Заменимые аминокислоты синтезируются животной клеткой из незаменимых аминокислот или других соединений.
Наличие аминогруппы с основными свойствами и карбоксильной группы с кислотными свойствами дает возможность аминокислотам соединяться в единую цепь за счет прочных ковалентных связей при отщеплении молекулы воды. Ковалентные связи называют пептидными, а последовательное соединение аминокислот в белковой молекуле пептидом: дипептид, трипептид, полипептид. Возможно огромное количество вариантов последовательных наборов 20-ти аминокислот в белковой молекуле.
Белки имеют первичную, вторичную, третичную и четвертичную структуру.
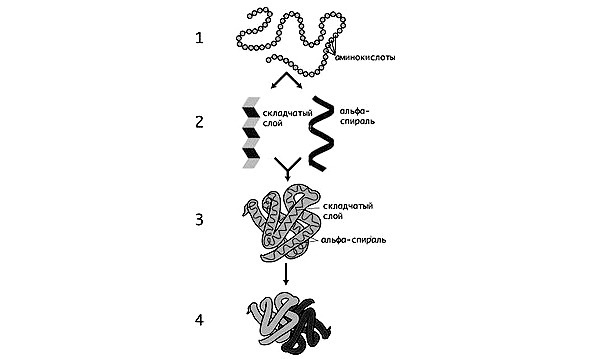
Первичная структура — это расположение аминокислот последовательно друг за другом в единую цепочку: — лизин — глутамин — валин — и т. д.
Вторичная структура представляет собой полипептидную цепь, закрученную ввиде спирали и имеющую слоисто-складчатое строение.
Третичная и четвертичная структура являются дальнейшим развитием белковой молекулы, которая усложняет пространственную укладку путем различного вида скручиваний. На уровнях третичной и четвертичной структур белки приобретают биологическую активность. Утрата белковой молекулой своей структурной организации называется денатурацией (происходит при изменении температуры, обезвоживании и пр.), а восстановление — ренатурацией. Главное условие полной ренатурации — сохранение первичной структуры белка.
Белки могут быть простыми и сложными. Простые белки состоят только из аминокислот, сложные белки имеют в своем составе другие органические соединения: нуклеиновые кислоты, углеводы, липиды, соединения фосфора, металлы. Соответственно их называют нуклеопротеиды, гликопротеиды, липопротеиды, фосфо- и металлопротеиды.
Функции белков в клетке
По разнообразию и значимости белки стоят на первом месте среди всех органических соединений. Им свойственны следующие функции:
1. Структурная — участие в строительстве клеточных мембран, хромосом, рибосом и других компонентов клетки.
2. Каталитическая. Белки-ферменты являются ускорителями внутриклеточных химических реакций, определяют их специфичность. Все известные ферменты делятся на две группы: простые (однокомпонентные), сложные (двухкомпонентные). Простые ферменты включают только белковую часть — апофермент. Сложные ферменты содержат белковую и небелковую части. Если небелковая часть легко отделяется и связывается с другими ферментами, ее называют коферментом. Ферменты могут иметь один или два активных центра. При наличии одного активного центра он связывается только с субстратом — веществом, на которое действует. В случаях двух активных центров один связывается с субстратом, другой с продуктом реакции. Все ферменты характеризуются специфичностью действия и саморегуляцией.
3. Энергетическая: при полном расщеплении 1г белка освобождается 17,6 кДж.
4. Сигнальная — белки, встроенные в поверхностные слои плазмалеммы (антигены), являются своеобразными «мишенями» для многих биологически активных веществ (гормоны).
5. Защитная — связанная с особой группой иммуноглобулинов, определяющих гуморальный иммунитет организма, кроме того многие белки образуют защитные покровы в виде чешуи, ногтей, копыт, волос и т. д.
6. Транспортная — с белками связан перенос ряда гормонов, а также кислорода (гемоглобин).
7. Двигательная — осуществляется сократительными (контрактильными) белками, с помощью которых происходит движение различных клеточных фибрилл (колебание жгутиков сперматозоидов, движение ресничек на поверхности клеток и др.)
Углеводы. Группа сложных органических соединений, в состав которых входят только атомы углерода, кислорода, водорода, азот отсутствует. Поскольку число атомов водорода в них в два раза превышает количество атомов кислорода, эти вещества названы углеводами.
Углеводы бывают простыми и сложными. Простые углеводы называют моносахаридами (мономеры). Сложные углеводы образованы несколькими мономерами и носят название полисахариды. Например, широко распространенные полисахариды крахмал, целлюлоза, гликоген в качестве мономера имеют глюкозу. Молекула целлюлозы образована цепочкой из нескольких сотен молекул глюкозы. Общая формула углеводов Сn (Н20) m. В зависимости от числа атомов углерода в молекуле моносахарида выделяют триозы (3 атома углерода), тетрозы (4 атома углерода), пентозы (5 атомов углерода), гексозы (6 атомов углерода). Наиболее важными моносахаридами, помимо глюкозы, являются рибоза и дезоксирибоза (пентозы), входящие в состав нуклеиновых кислот. Растительные клетки содержат значительно больше углеводов, чем животные.
Функции углеводов. Главная роль углеводов — энергетическая. При окислении 1 г углеводов выделяется 17,6 кДж. Углеводы выполняют также структурную роль, входя в состав плазмалемм клеток (гликокаликс) и клеточных оболочек (целлюлоза).
Липиды представляют органические вещества нерастворимые в воде, но хорошо растворимые в эфире, бензине, ацетоне и др. Сами липиды могут являться растворителем для некоторых веществ, например, витаминов А, Е. По химическому составу липиды разнообразны и включают жирные кислоты, аминоспирты, аминокислоты, фосфорную кислоту. Между этими соединениями образуются различные виды химических связей. Все липиды делят на две большие группы: нейтральные жиры и фосфолипиды. Нейтральные липиды являются производными высших жирных кислот и трехатомного спирта глицерина. Обычно количество липидов в клетках невелико, всего 1,0—1,3%, но в некоторых специализированных клетках они составляют основную массу цитоплазмы (жировые клетки, отдельные виды яйцеклеток).
Главные функции липидов: структурная и энергетическая. Липиды входят в состав клеточных мембран (фосфолипиды). При расщеплении 1 г липидов выделяется 38,9 кДж энергии.
Строение остальных органических соединений клетки — нуклеиновых кислот, АТФ освещено в соответствующих главах.
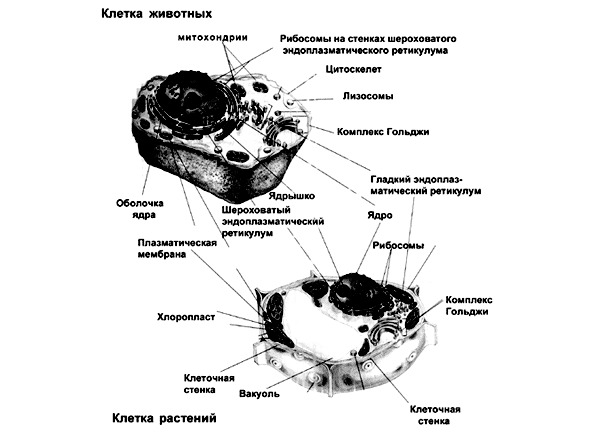
Структурная организация клетки
Основными структурными компонентами клетки являются клеточные мембраны, ядро, цитоплазма с цитоскелетом, органеллы и включения.
І. Клеточная мембрана или плазмалемма представляет собой тонкую биологическую пленку, ограничивающую клетку. Она обеспечивает разделение двух фаз: внеклеточной со случайным набором ионов и молекул и внутриклеточной со строго упорядоченным их составом. Для поддержания таких концентрационных градиентов мембрана должна удовлетворять одному абсолютному требованию — необходима ее полная замкнутость. Поэтому все известные биологические мембраны образуют замкнутые пространства — компартменты. Другое важное свойство плазмалеммы — асcимметричность: ее внутренняя и наружная поверхность должны функционировать по-разному. В противном случае молекулы и ионы, вносимые в одном месте, будут столь же быстро выноситься в другом. Таким образом, главная функция клеточной мембраны — обеспечить поступление в клетку веществ и сохранить постоянство ее состава, то есть клеточный гомеостаз.
Основу плазмалеммы составляет двойной слой липидов, расположенных перпендикулярно поверхности. Липиды представлены фосфолипидами и холестеролом. Именно они обеспечивают структурную целостность мембраны. Оба вида липидов амфипатические: один конец молекулы — «головка» — полярный гидрофильный, другой конец — «двойной хвост» — неполярный гидрофобный. Если гидрофильную головку отделить от молекулы, она растворится в воде. Гидрофобный хвост, подобно растительному маслу, в воде не растворим. Гидрофильные головки липидов обращены кнаружи, а гидрофобные концы спрятаны вовнутрь (рис. 4).
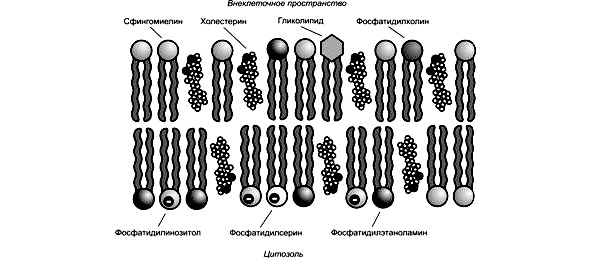
Липидный бислой плазмалеммы прикрыт с обеих сторон белками, которые подразделяются на два класса. Первый класс — трансмембранные белки. Определенная часть их молекулы встроена в двойной липидный слой и пронизывает его насквозь.
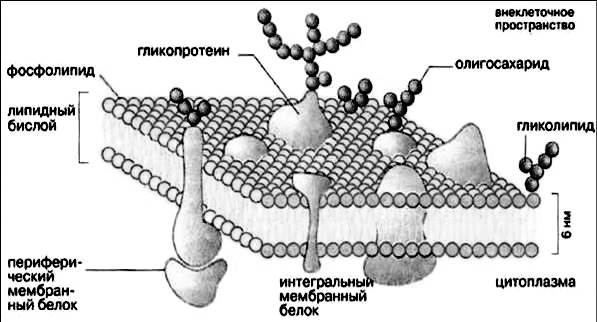
В мембране возникают белковые поры. Второй класс составляют периферические белки, которые не встроены в липиды, а находятся на поверхности. Здесь они вступают в связь с определенными трансмембранными белками. Тем не менее, их можно отделить от мембраны и растворить в воде, в то время как трансмембранные белки не отделимы от липидов и не растворимы в водной среде. Периферические белки сконцентрированы на внутренней и наружной поверхности плазмалеммы. На наружной поверхности к ним примыкают углеводы, формирующие тонкое покрытие клетки — гликокаликс (см. рис. 5).
Функции клеточной мембраны
Клеточная мембрана выполняет две основные функции:
а) межклеточные взаимодействия; б) транспортные процессы.
Обе эти функции во многом определяются белками или гликопротеинами (сложные комплексы белков и углеводов) клеточной мембраны.
Межклеточные взаимодействия. Клетка воспринимает и трансформирует сигналы двух родов: из внешней среды и внутренней среды организма. Раздражители из внешней среды могут иметь различную природу: физическую (кванты света), химическую (вкусовые молекулы), механические (сжатие или растяжение). Из внутренней среды поступают сигналы преимущественно информационного характера, например, гормоны, нейромедиаторы.
Межклеточные взаимодействия реализуются по принципу сигнал — ответ. Сигнал воздействует на специфический клеточный рецептор, который обычно представлен особым поверхностным белком или гликопротеином. Ответ состоит в адекватной реакции клетки.
Транспортные процессы. Перенос веществ через клеточную мембрану может осуществляться в виде пассивного транспорта, не требующего затрат энергии и активного транспорта, сопряженного с потреблением энергии.
Пассивный транспорт связан с движением по градиенту концентрации. Он может происходить в обоих направлениях: из клетки в среду и из среды в клетку. Так движутся небольшие полярные (СО2, Н20) и неполярные (02, N2) молекулы. Движение в одном направлении, в клетку, происходит путем облегченной диффузии. В ней участвуют специальные трансмембранные белки, которые называются белки-переносчики. Такие специальные белки существуют для глюкозы, аминокислот, фосфатов и др.
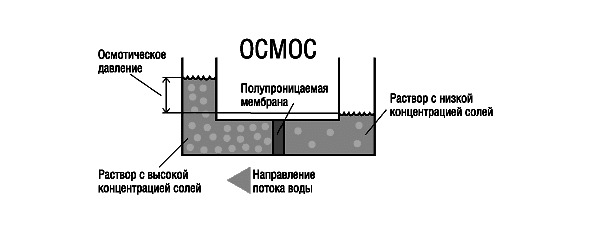
Осмос — перенос вещества из одного раствора в другой через мембрану. Мембраны, проницаемые не для всех веществ, а лишь для определенных, называют полупроницаемыми. Осмос играет важную роль во многих биологических процессах. Мембрана, окружающая нормальную клетку крови, проницаема лишь для молекул воды, кислорода, растворенных в крови питательных веществ и продуктов клеточной жизнедеятельности. Она непроницаема для больших белковых молекул, находящихся в растворенном состоянии внутри клетки. Поэтому белки, столь важные для биологических процессов, остаются внутри клетки.
Активный транспорт происходит против градиента концентрации и потому является энергозависимым. Он происходит при участии ферментов АТФаз. Наиболее известен натрий-калиевый насос. Его функция выкачивать из клетки Na+ в обмен на К+. Тем самым, насос обеспечивает связанный транспорт множества органических и неорганических молекул. Натрий-калиевый насос — это особый трансмембранный белок. С внутренней стороны клетки к нему поступает Na+ и АТФаза, с наружной К+. Посредством изменения формы белка из клетки активно выкачивается Na+ и закачивается К+.
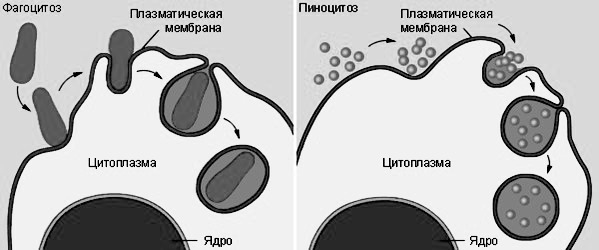
Транспортные клеточные процессы могут осуществляться также путем эндоцитоза и экзоцитоза. При эндоцитозе клетки поглощают макромолекулы и оформленные частицы, окружая их небольшим участком клеточной мембраны. Последняя впячивается вовнутрь клетки, образует везикулы (пузырьки). Если везикулы мелкие и содержат внеклеточную жидкость, процесс называется пиноцитоз. Если они содержат крупные компактные частицы, то формируются фагосомы, а явление известно как фагоцитоз (рис. 6).
Экзоцитоз — это выброс веществ из клетки в виде гранул секрета или вакуолей с клеточной жидкостью.
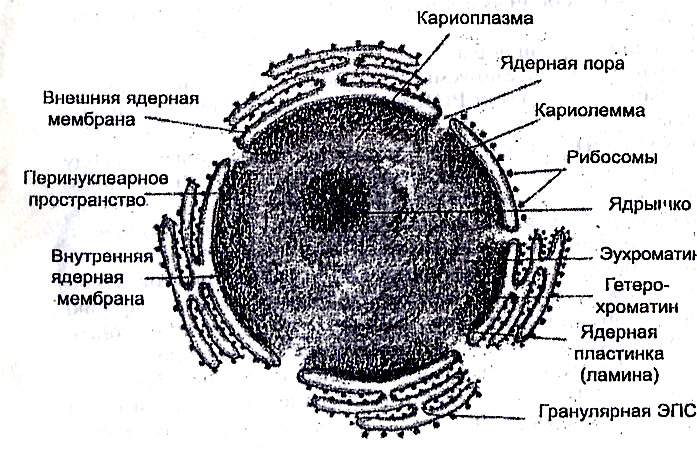
ІІ. Ядро — центральный аппарат клетки, с которым связано хранение и передача генетической информации, обмен веществ, движение и размножение. Форма ядра чаще округлая или вытянутая, реже дольчатая. От цитоплазмы его отделяет кариолемма (ядерная оболочка). Она состоит из наружной и внутренней ядерных мембран, разделенных бесструктурным веществом. Мембраны имеют поры, обеспечивающие избирательную связь с цитоплазмой. Общее их количество в клетке 3—4 тысячи. Каждая пора встроена в крупную дисковидную структуру, называемую поровый комплекс ядерной оболочки. Заполнено ядро гомогенной массой нуклеоплазмой. В ее состав входят нуклеиновые кислоты и белки.
Комплекс ядерной ДНК со структурными белками (гистонами и негистоновыми белками) называют хроматином. На цитологических препаратах хроматин имеет вид глыбок различной величины и формы. Выделяют эухроматин, генетически активную структуру и гетерохроматин, генетически инертную составляющую.
В период деления клетки в ядре выявляются митотические хромосомы. Они выглядят как короткие палочковидные тельца, обладающие особой индивидуальностью и функцией. Важным компонентом ядра является одно или несколько ядрышек. Это мелкие круглые тельца с высоким содержанием РНК и белка. Ядрышковая РНК участвует в синтезе р-РНК и образовании рибосом (рис. 7).
Генетический аппарат ядра клетки вырабатывает определенные белки-инвенторы, которые регулируют проницаемость клеточной мембраны. Важным элементом старения клетки служит прекращение синтеза инвенторов.
ІІІ. Цитоплазма объединяет все живое вещество клетки, за исключением ядра и ограничивающих клетку мембран. Гомогенная бесструктурная масса цитоплазмы получила название гиалоплазмы (греч. hyalos — стекло и плазма). В ней во взвешенном состоянии находятся органеллы и включения. Агрегатное состояние цитоплазмы бывает жидкое — золь и вязкое — гель. Основу цитоплазмы формирует цитоскелет клетки.
Цитоскелет — сложная сеть микротрубочек и белковых филаментов (нитей). Микротрубочки имеют в диаметре 10—25 нм, содержат белок тубулин. С их помощью происходит транспорт веществ и движение органелл. Микрофиламенты тоньше, в диаметре 4—7 нм, включают белки актин, миозин и выполняют сократительную функцию.
Движение цитоплазмы называется циклоз. Это перемещения самой цитоплазмы или ее структур без изменения формы клетки, например, при клеточной регенерации. Тем самым циклоз отличается от цитокинеза при митозе или амебоидного движения при фагоцитозе. Цитоплазматические движения обеспечивают элементы цитоскелета, они во многом зависят от агрегатного состояния содержимого клетки золь — гель.
Органеллы и включения цитоплазмы
В цитоплазме находятся органеллы и включения. Органеллы — это постоянные высокодифференцированные внутриклеточные образования, выполняющие определенные функции. Они имеют мембранный и немембранный принцип строения.
Классификация органелл
1. Органеллы общего значения: эндоплазматический ретикулум, рибосомы, митохондрии, комплекс Гольджи, лизосомы, центросомы, пластиды.
2. Органеллы специального значения: реснички, жгутики, миофибриллы, нейрофибриллы.
Органеллы общего значения
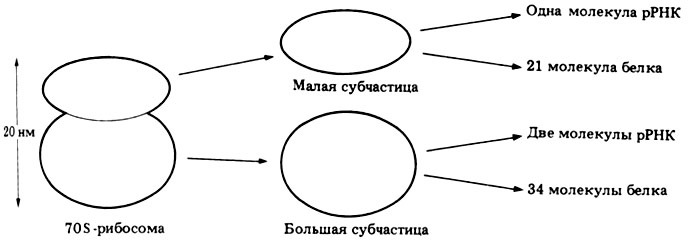
Эндоплазматический ретикулум (ЭР). Разветвленная внутриклеточная структура, представленная системой субмикроскопических канальцев с расширениями-цистернами. Они ограничены мембранами, несущими многочисленные ферменты. Мембраны ЭР составляют более половины массы всех мембран клетки. Существует два типа ЭР. Гранулярный ЭР, мембраны которого содержат рибосомы. Рибосомы — это ультрамикроскопические, сферические гранулы, состоящие из двух половинок — большой и малой субъединиц, а также рибосомальной РНК. Главное назначение их — участие в синтезе белка (рис. 8).
Гладкий ЭР несет мембраны, лишенные рибосом. Здесь происходит синтез липидов и углеводов. ЭР объединен с ядром клетки, поскольку наружная мембрана ядра непосредственно переходит в мембраны ЭР. Гладкий и гранулярный ЭР связаны друг с другом, но отличаются по составу содержащихся в них белков. Все белки, подлежащие выведению из клетки, проходят путь от рибосом в полость ЭР, оттуда в аппарат Гольджи, а затем в секреторные везикулы (пузырьки), которые выводятся из клетки. Белки, предназначенные для внутриклеточных нужд, используются митохондриями или другими органеллами и никогда не поступают в ЭР (рис. 9).
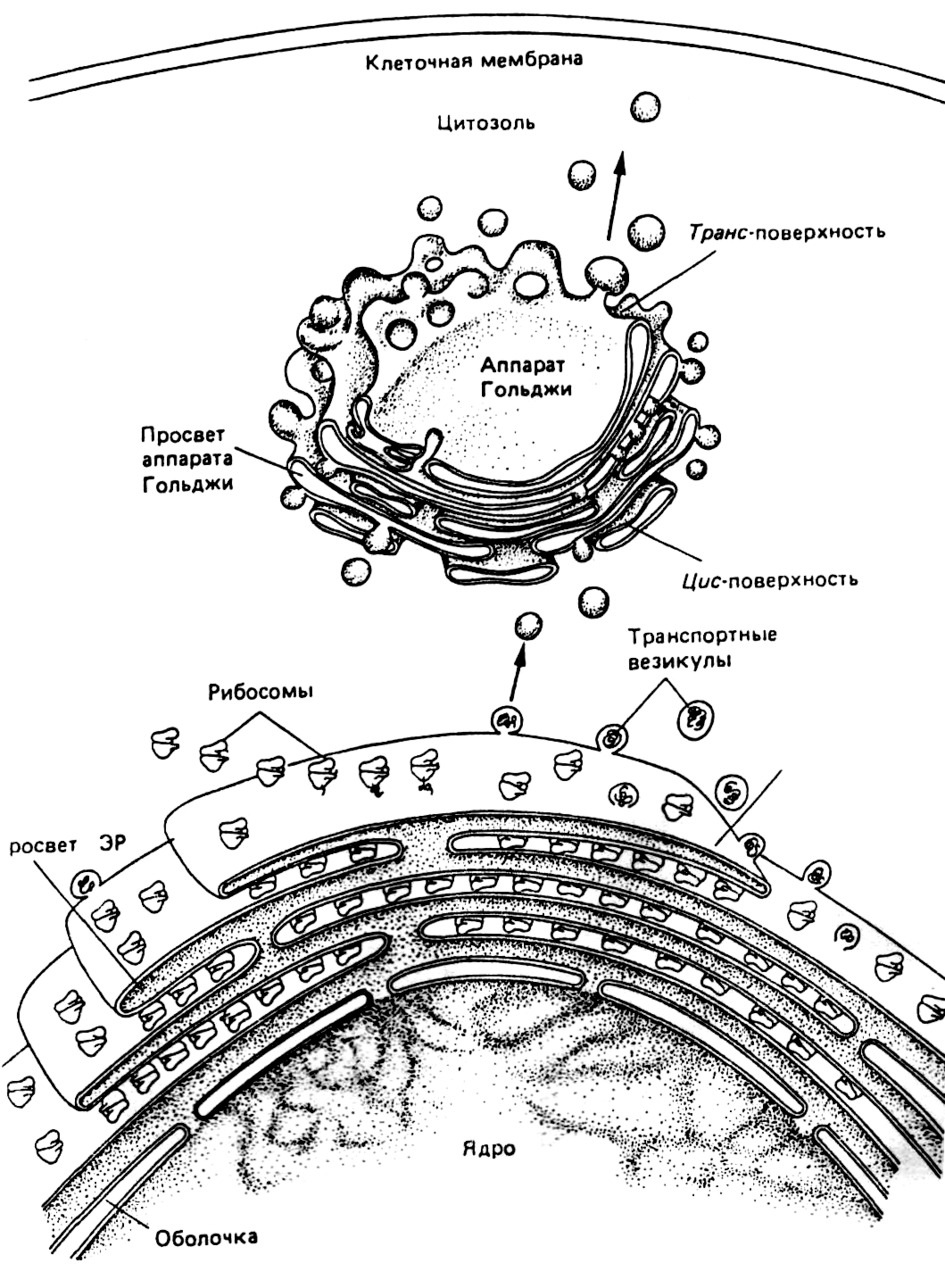
Липопротеиды, входящие в состав мембран ЭР, очевидно, аналогичны тем, что входят в состав наружной клеточной мембраны. Обе мембраны могут соединяться и тогда ЭР открывается наружу клетки, в частности, это происходит в случаях клеточной секреции. С другой стороны, удается рассмотреть тот момент, когда субстанции, проникающие в клетку, появляются в ЭР, при этом они, как правило, направляются к лизосомам. Следовательно, ЭР оберегает клетку от вторжения в нее инородных субстанций. В то же время, ЭР может являться пристанищем для некоторых вирусов, в частности ретровирусов. В условиях патологии наблюдается два вида морфологических изменений — гиперплазия и атрофия эндоплазматического ретикулума.
С эндоплазматическим ретикулумом нередко связаны пероксисомы или микротельца. Они имеют округлые очертания, окружены одинарной мембраной, диаметр их не более 1,5 мкм. Содержат фермент пероксидазу, катализирующий расщепление пероксида водорода на воду и кислород. Уменьшение числа пероксисом и снижение синтеза их ферментов наблюдается в печени при воспалении, а также при опухолевом росте.
Митохондрии. В конце позапрошлого века в цитоплазме различных клеток были выявлены нитевидные и гранулярные структуры. Ученый Бенда назвал их митохондрии, от греческого «митос» — нить и «хондрос» — зерно. Величина этих органелл 0,5—5,0 мкм. Количество может варьировать от нескольких единиц до десятков тысяч. Как показала электронная микроскопия, митохондрии имеют наружную и внутреннюю мембрану. Наружная мембрана напоминает сито, проницаемое для небольших молекул массой менее 10000 Да. Промежуток между наружной и внутренней мембраной называют межмембранным пространством. Внутренняя мембрана образует многочисленные складки, кристы, в виде гребней, вдающихся во внутреннюю полость, называемую матрикс (рис. 10). Кристы значительно увеличивают общую площадь митохондриальных мембран, что определяет высокую функциональную активность.
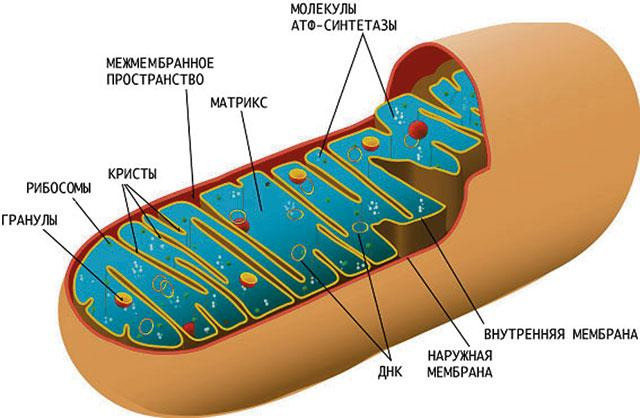
На кристах содержатся комплексы дыхательных ферментов, необходимые для окислительного фосфорилирования. Результатом его является образование АТФ и выделение большого количества энергии, необходимой для жизнедеятельности клеток. Митохондрии содержат цитоплазматическую ДНК, отличную от ДНК ядра.
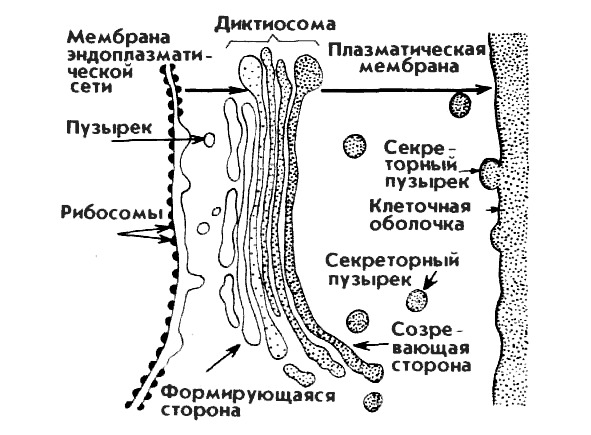
Комплекс Гольджи. Указанная органелла названа именем итальянца Карелло Гольджи, открывшего ее в 1896 г. В световом микроскопе структура имеет вид сетки, лежащей вблизи ядра. По данным электронной микроскопии комплекс состоит из диктиосом. Каждая диктиосома представляет стопку плоских мешочков-цистерн диаметром около 7 мкм. Число цистерн в одной диктиосоме 5—7. От краев цистерн отделяются микропузырьки. Вся структура имеет полярный характер, поскольку в диктиосоме два полюса — формирующий у основания и секретирующий у вершины. Основная функция комплекса Гольджи заключается в накоплении и конденсации продуктов, синтезируемых эндоплазматическим ретикулумом (рис. 11). Образно говоря, здесь происходит упаковка сложных химических соединений в виде пузырьков, гранул, зерен, которые затем выводятся из клетки. Величина аппарата Гольджи связана с синтетической активностью клетки и обусловлена либо уровнем наружной секреции, например, в печени или поджелудочной железе, либо интенсивностью синтеза, необходимой для жизнедеятельности самой клетки, например, в нейронах.
Лизосомы. В названии данной органеллы биохимик Де Дюву объединил два греческих слова: «лизис» — растворение и «сома» — тело. Лизосомы представляют сферические частицы размерами 0,5—2,0 мкм (рис. 12). Они имеют плотную липопротеиновую мембрану и содержат большой набор гидролитических ферментов, необходимых для процессов внутриклеточного пищеварения. На это указывает высокое содержание лизосом в клетках-фагоцитах, а также присутствие лизосомальных ферментов во внутриклеточных пищеварительных вакуолях. Другой важной функцией лизосом является аутолиз — посмертное растворение структурных компонентов клетки под действием все тех же ферментов лизосом. Как выразился Де Дюву — это «маленькие могильщики», освобождающие ткани от мертвых клеток.
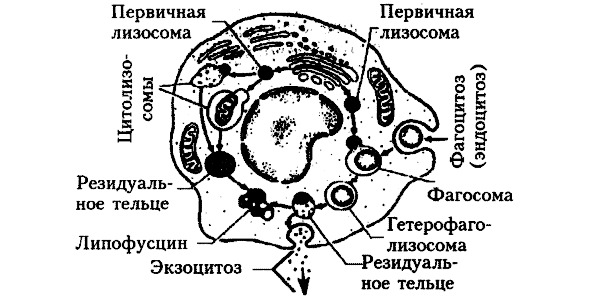
В лизосомах могут отсутствовать некоторые энзимы, необходимые для нормального метаболизма клеток. Энзимопатия или дисметаболическая болезнь имеет врожденный характер и наследуется по аутосомно-рецессивному типу. Дефицит энзимов наблюдается наиболее часто при гликогенозах (болезнь Помпе, болезнь Гирке), липидозах (недостаточность липаз адипозоцитов), гепатозах (болезнь Дабина-Джонсона). Эти состояния иногда называют «болезнями накопления». В реальной действительности речь идет не об избыточном образовании различных субстанций, а о замедлении или остановке разрушения их метаболитов при нормальном синтезе.
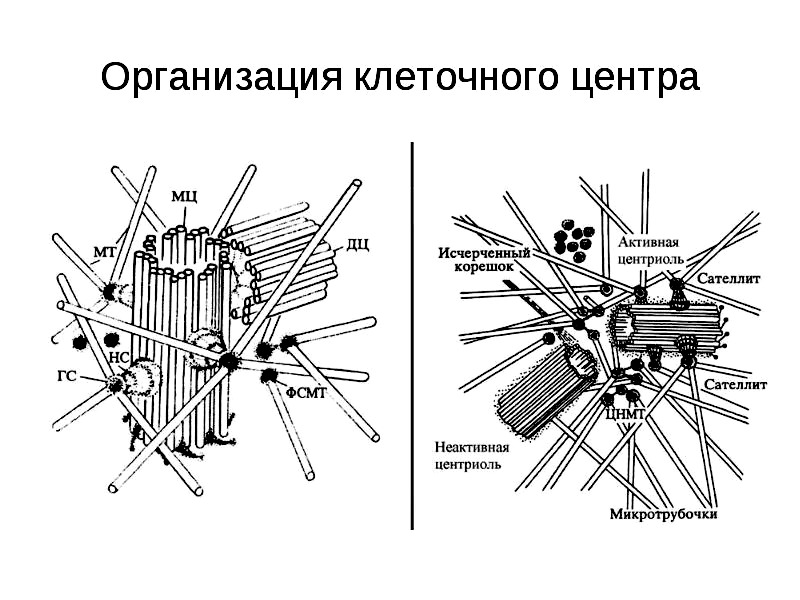
Центросома. Открыл эту структуру голландский ученый Ван-Бенеден, в 1876 г., но название предложил цитолог Бовери, опять-таки, используя греческую терминологию: «центрум» — центр и «сома» — тело. Типичная центросома представлена двумя центриолями, соединенными перемычкой — центродесмосомой и окруженной «лучистой» сферой — астросферой. При электронной микроскопии центриоли имеют вид цилиндра, стенки каждого образованы микротрубочками. Центросома обеспечивает процесс митоза, формируя митотическии аппарат клетки (рис. 13).
Органеллы специального значения
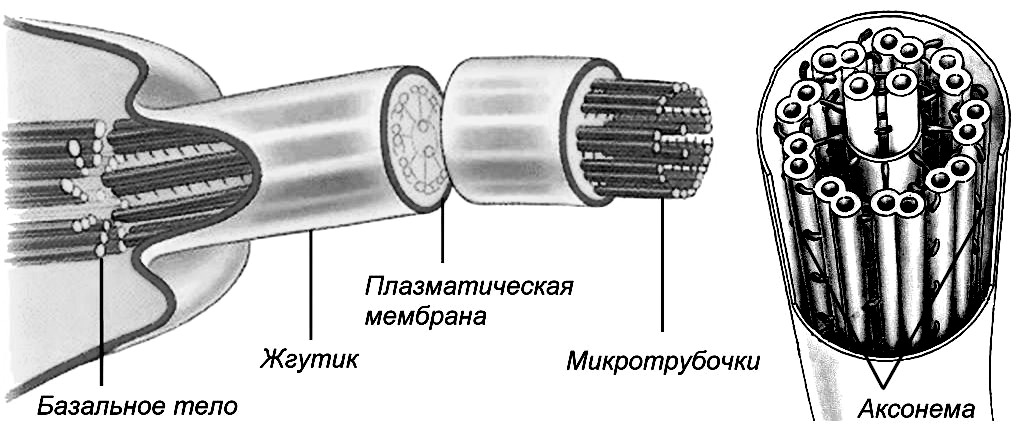
Реснички и жгутики. Эти органоиды встречаются у одноклеточных организмов (бактерии, простейшие) и у клеток в составе тканей (клетки эпителия трахеи). Реснички и жгутики представляют тончайшие подвижные выросты цитоплазмы, в которых находятся ультратонкие фибриллы, содержащие белок с сократительной функцией (рис. 14). Они отражают элементы движения. Существуют генетические аномалии строения ресничек. Например, врожденный синдром неподвижных ресничек (синдром Картагенера) характеризуется тем, что реснички покровного эпителия дыхательных путей и слизистой оболочки среднего уха неподвижны или малоподвижны. Поэтому мукоцилиарный транспорт резко ослаблен или отсутствует, что ведет к хроническому воспалению дыхательных путей и среднего уха. У таких больных неподвижны также сперматозоиды, так как их хвост эквивалентен ресничкам.
Миофибриллы. Характерны для мышечных клеток. Состоят из элементарных ультратонких нитей протофибрилл. Описано два типа миофибрилл — гладкие и поперечнополосатые. Последние содержат систему светлых (изотропных) и темных (анизотропных) дисков. Выполняют сократительную функцию.
Нейрофибриллы. Являются обязательным компонентом многих нервных клеток и их отростков. Они могут быть очень тонкими гладкими однородными нитями (нейропротофибриллы) или более толстыми трубчатыми элементам, имеют опорное значение в связи с наличием в нервных клетках отростков большой протяженности.
Включения — непостоянные структурные компоненты клетки, возникающие в результате внутриклеточного метаболизма или других процессов жизнедеятельности клетки. Исходя из особенностей субстрата, различают жидкие включения — вакуоли и плотные включения — гранулы, кристаллы. Обе группы включений могут нести на своей поверхности ограничивающие их тонкие липопротеиновые мембраны.
В функциональном отношении все включения подразделяются на три группы: трофические, секреторные и специальные.
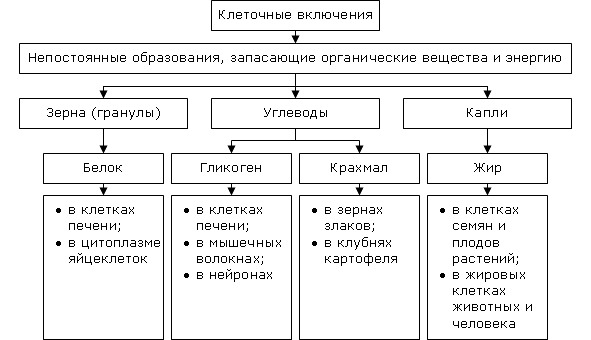
Трофические включения отражают повседневный метаболизм клетки. Они представлены гранулами гликогена, белковыми зернами, каплями жира. В ходе внутриклеточного обмена включения этой группы утилизируются и входят в состав цитоплазмы. Особую группу трофических включений составляют продукты экскреции (распада), подлежащие выведению из клетки: желчные пигменты, мочевина и др.
Секреторные включения характерны, в основном, для железистых клеток. Значение их очень велико, так как сюда относятся некоторые ферменты и гормоны.
Специальные включения присутствуют в высокоспециализированных клетках. К этой группе относят гранулы пигмента меланина, плотно заполняющего цитоплазму меланоцитов — особых клеток кожи с защитной функцией. Диффузное состояние специальных включений выражает гемоглобин, находящиеся в клетках крови — эритроцитах. Включения могут характеризовать патологическое состояние клетки и быть предвестниками ее гибели (появление слизистых конгломератов в клетках кости или хряща).
Таким образом, приведенные выше данные указывают, что внутриклеточные компоненты имеют мембранный и немембранный принцип организации. Мембранные структуры можно подразделить на одномембранные и двумембранные. Одномембранные компоненты имеют вид каналов, цистерн, пузырьков, ограниченных одной мембраной и тесно взаимосвязанных. Эти пузырьки иногда называют «вакуолярной системой», хотя данное определение недостаточно точно. Сюда можно отнести: а) эндоплазматический ретикулум; б) комплекс Гольджи; в) лизосомы; г) вакуоли у растительных клеток и некоторых простейших.
Двумембранные компоненты — это митохондрии и пластиды. Наружная мембрана их всегда гладкая, внутренняя образует выросты, имеющие важное функциональное значение. Систему двойных мембран имеет также ядро — центральный аппарат клетки. Ядерные мембраны содержат поры.
Немембранные структуры клетки немногочисленны и в той или иной мере связаны с системой мембран. В число их входят: а) рибосомы, состоящие из двух субъединиц; б) центросома, локализованная вблизи ядра: в) органеллы движения клеток — жгутики; реснички, миофибриллы; г) разнообразные клеточные включения.
Обмен веществ и превращение энергии в клетке
Клетка — это открытая система. В ней непрерывно происходит обмен веществ. Он включает поступление в клетку неорганических и органических соединений, их превращения и выведение из клетки. Тем самым, обмен веществ имеет две стороны: пластический обмен (ассимиляция) и энергетический обмен (диссимиляция).
Универсальный источник энергии клетки — АТФ
Для того чтобы существовать и выполнять определенные функции клетка нуждается в энергии. Энергия, приобретаемая клеткой, сохраняется в полезной форме, главным образом в виде молекул аденозинтрифосфата — АТФ (аденозинтрифосфорная кислота). Молекула АТФ является нуклеотидом, так как состоит из азотистого основания — аденина, сахара рибозы и трех фосфатных групп (остатки фосфорной кислоты). АТФ — это макроэргическое соединение поскольку в двух фосфатных связях накапливается большое количество энергии. Химические связи, которыми соединены молекулы фосфорной кислоты неустойчивы. Под действием фермента АТФ-азы в ходе гидролиза (присоединения воды) один богатый энергией остаток фосфорной кислоты отщепляется от молекулы АТФ с образованием аденозиндифосфата АДФ и выделением энергии в количестве около 40 кДж/моль. Указанный процесс называется дефосфорилированием. Обратное явление, переход АДФ в АТФ путем присоединения неорганического фосфата — фосфорилированием.
Пластический обмен. Биосинтез белка
Информационное обеспечение процессов синтеза. Живая клетка содержит тысячи различных белков. Более того, каждый вид клеток имеет свои специфические белки. Эти белки клетка обязана синтезировать, передать потомкам и воспроизвести затем в ряду поколений. Следовательно, любая клетка должна обладать биологической памятью. Биологическая память — это хранение и передача клеткой генетической информации. Обеспечивают ее нуклеиновые кислоты.
Нуклеиновые кислоты
Нуклеиновые кислоты открыты еще в позапрошлом веке швейцарским ученым Ф. Мишером (1868 г.). Первые экспериментальные данные о значении нуклеиновых кислот в механизмах наследственности были получены группой микробиологов, возглавляемой О. Эвери в 1944 г. Ученые работали с двумя близкими видами бактерий, вызывающих воспаление легких. Один вид имел хорошо выраженную полисахаридную капсулу, другой нет. Выделив нуклеиновую кислоту, ДНК от бактерий с капсулами и обработав ею бескапсульные микроорганизмы, исследователи получили в потомстве последних как капсульные, так и бескапсульные формы. Это указывало на хранение информации о наличии капсулы именно в молекуле ДНК. В дальнейшем было представлено огромное количество фактов, подтверждающих это открытие.
Существует два класса нуклеиновых кислот:
ДНК — дезоксирибонуклеиновые кислоты. РНК — рибонуклеиновые кислоты.
ДНК локализована в ядрах, митохондриях и пластидах клеток. РНК входит в состав ядрышек, рибосом и присутствует в цитоплазме клеток. По своей химической структуре ДНК и РНК — это крупные молекулы биополимеров. Они складываются из мономеров — нуклеотидов. Каждый нуклеотид включает три компонента: полисахарид, азотистое основание, фосфат (остаток фосфорной кислоты) (рис. 15).
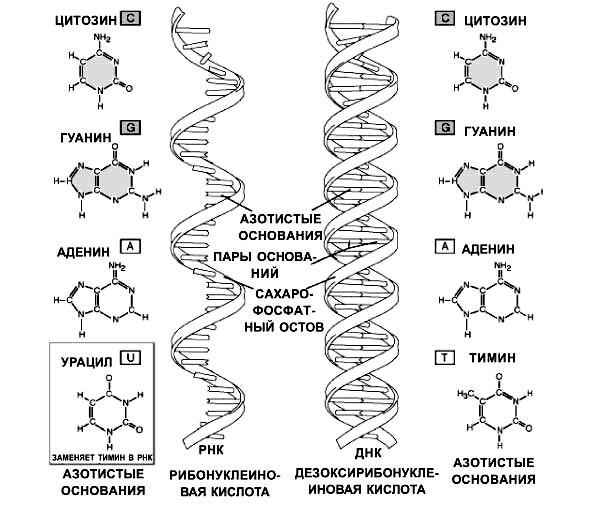
В состав нуклеотида молекулы ДНК входят углевод — дезоксирибоза (пентоза), остаток фосфорной кислоты и одно из четырех имеющихся азотистых оснований. Азотистые основания пуриновые — аденин (А), гуанин (Г) и пиримидиновые — цитозин (Ц), тимин (Т).
По правилу Чаргаффа общее количество пуриновых оснований в молекуле ДНК равно количеству пиримидиновых оснований: пуриновые А + Г = Ц + Т пиримидиновые
В 1953 г. Д. Уотсон и Ф. Крик, используя кристаллографический анализ, построили модель пространственной структуры молекулы ДНК, за что были удостоены Нобелевской премии. Основные положения этой модели следующие:
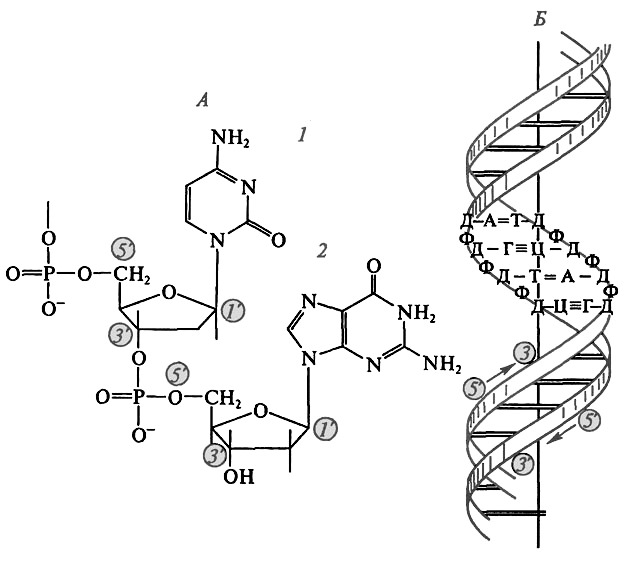
1. ДНК образована двумя полинуклеотидными цепями, при этом соединение нуклеотидов в цепь обеспечивается ферментом ДНК-полимеразой.
2. Цепи правозакручены спирально вокруг общей оси, один виток включает 10 пар нуклеотидов.
3. Фосфатные группировки находятся снаружи спирали, а азотистые основания внутри.
4. Цепи антипараллельны, то есть последовательность атомов одной цепи противоположна таковой в другой.
5. Цепи удерживаются водородными связями между азотистыми основаниями по принципу комплементарносги. Аденин соединяется двумя связями с тимином, гуанин тремя связями с цитозином (рис. 16).
Открытие явления комплементарности — крупнейшее достижение биологии XX века. Следует заметить, что водородные связи слабые и нарушаются при изменении pH клетки или нагревании до 100°. Исходя из сказанного, различают первичную и вторичную структуру ДНК.
Первичная структура — это линейная последовательность нуклеотидов в антипараллельных цепях.
Вторичная структура — сближение азотистых оснований по принципу комплементарности, их спирализация, что придает молекуле ДНК высокую компактность. Как показал академик Белозерский, ДНК, выделенная из разных тканей одного организма, имеет одинаковый нуклеотидный состав, а соотношение пар азотистых оснований является строгим видовым признаком.
Функции ДНК
1) аутосинтетическая — репликация (синтез ДНК) в S-периоде интерфазы клеточного цикла;
2) гетеросинтетическая — участие в синтезе белка, хранении и передачи генетической информации.
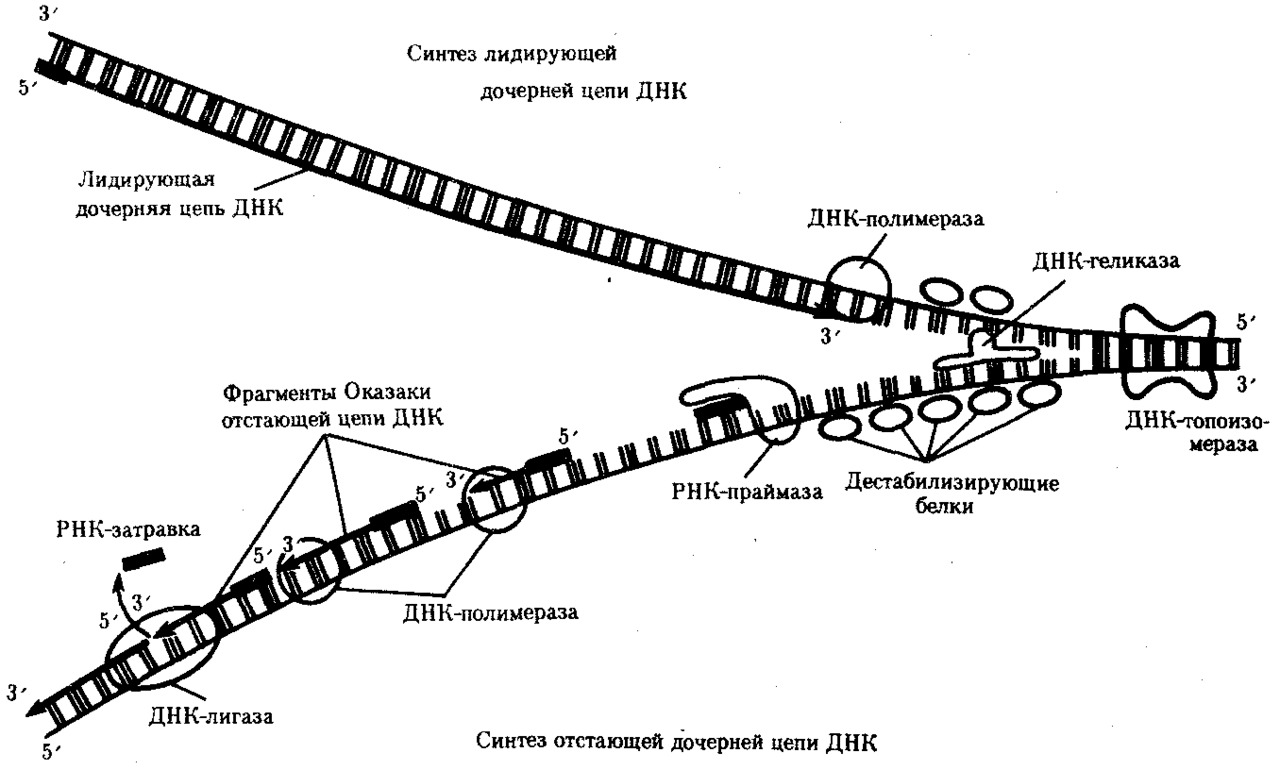
При репликации происходит следующее:
а) разрыв водородных связей между двумя полинуклеотндными цепями и их расхождение;
б) деспирализация полинуклеотндных цепей;
в) синтез новых цепей вдоль каждой из разделившихся, но правилу комплементарности с точным сохранением их структуры.
Описанная репликация называется полуконсерватнвной, поскольку одна из цепей новой молекулы ДНК является старой, а другая вновь синтезированной. Репликация протекает при участии ферментов ДНК-полимераз (рис.17).
ДНК — это единственная макромолекула клетки, которая способна устранять повреждения, возникающие в ее структуре, что может приводить к мутациям. Более того, в ней закодирована информация о механизмах самых разнообразных репарационных процессов.
РНК
Общий принцип структурной организации нуклеиновых кислот сохранен, но имеется ряд особенностей. Сахарид представлен рибозой (содержит на один атом кислорода больше. чем дезоксирибоза). Азотистое основание тимин заменено на урацил (Т-У). В молекулах РНК только одна полинуклеотидная цепь, то есть они одноцепочечные.
Выделяют три разновидности РНК
1) и-РНК, информационная;
2) т-РНК, транспортная;
3) р-РНК, рибосомальная.
Молекула и-РНК синтезируется на молекуле ДНК в ядре клетки и затем переносится в цитоплазму; т-РНК присутствует в цитоплазме и осуществляет транспорт аминокислот; р-РНК находится в рибосомах.
Функция РНК — участие в синтезе белка. Сколь бы замечательны не были белки, они лишены одного очень важного свойства. Белок в составе клетки не может точно воспроизвести сам себя на протяжении ряда клеточных делений. Точное копирование белковых молекул происходит с помощью генетического кода. Под ним понимают систему записи в молекулах ДНК генетической информации о строении белковой молекулы. Белок состоит из аминокислот, которых всего 20. Аминокислоты в белковой молекуле расположены в линейном порядке, подобно нуклеотидам молекулы ДНК. Строгая последовательность аминокислот определяет первичную структуру белковой молекулы, ее специфичность. Так, в составе белка гемоглобина 300 аминокислот. Достаточно изменить лишь одну аминокислоту в общей цепи, чтобы свойства гемоглобина нарушились. Последовательность аминокислот в белке определяется последовательностью нуклеотидов в молекуле ДНК, его генетическим кодом.
Принципы генетического кода
1. Код триплетный: три нуклеотида, расположенные рядом в строгой последовательности, соответствуют определенной аминокислоте. Триплет нуклеотидов называют кодоном. Например, аминокислоту валин кодирует кодон ЦАТ.
2. Код специфичен. Нет случаев, когда один и тот же кодон соответствовал бы нескольким аминокислотам.
3. Код вырожденный или избыточный, поскольку одной аминокислоте может соответствовать несколько триплетов. Валин кодирует не только кодон ЦАТ, но и ЦАЦ. Это свойство имеет существенное значение для повышения надежности системы хранения генетической информации.
4. Код однонаправленный. При синтезе белка считывание информации идет от первого основания к третьему в одном направлении.
5. Код неперекрывающийся. Это значит, нет таких оснований, которые принадлежали бы одновременно двум кодонам. Триплет ограничен как бы «рамкой» считывания, которая перемещается сразу на три основания: УГГ ЦАА.
6. Код является универсальным для всех живых существ.
Синтез белка в клетке
Синтез белка включает несколько уровней (рис. 18)
1. Претранскрипционный. 2. Транскрипционный. 3. Транспортный.
4. Трансляционный. 5. Посттрансляционный.
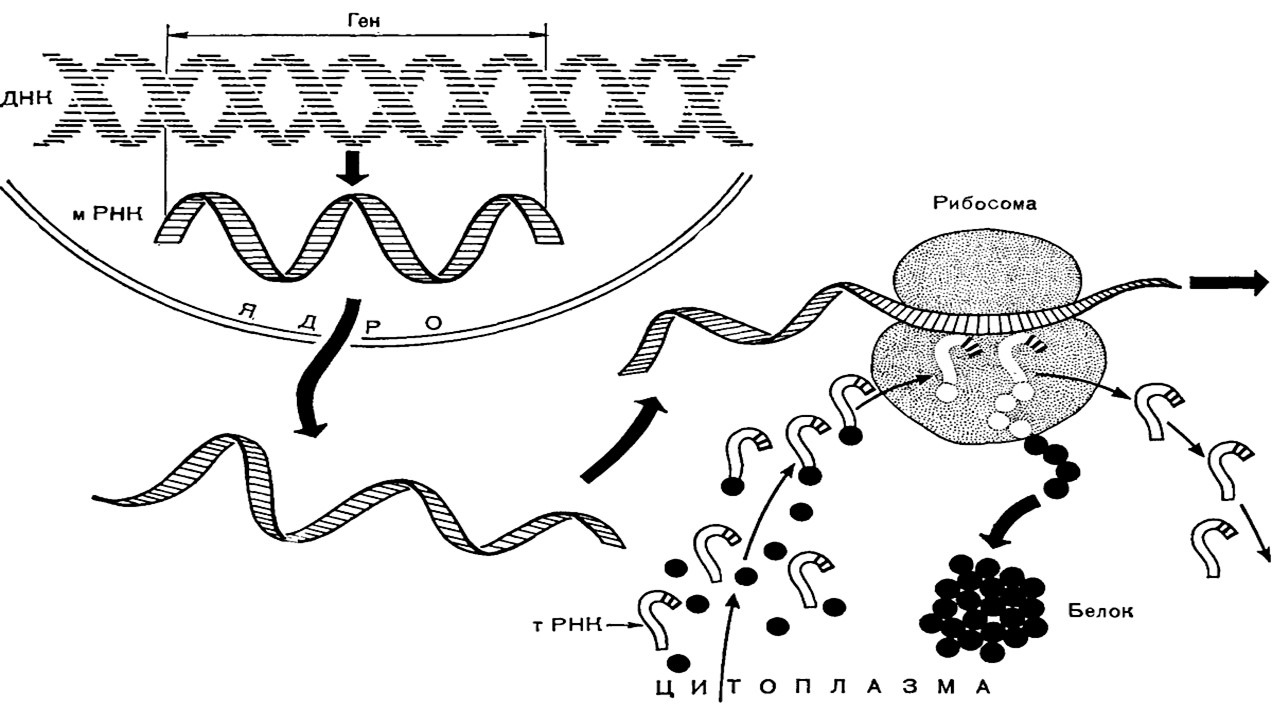
Претранскрипционный уровень. Это стартовый этап синтеза, во время которого происходит активация молекулы ДНК с помощью специальных белков.
Транскрипционный уровень. Транскрипцией называется синтез и-РНК на молекуле ДНК с последовательностью нуклеотидов комплементарной молекуле ДНК. Матрицей для и-РНК служит только одна из двух имеющихся цепей ДНК, то есть синтез носит полуконсервативный характер. Процесс контролируется ферментами РНК-полимеразами.
В молекуле ДНК выделяется определенный фрагмент — промотор как начальный участок синтеза. За ним следуют информационные нуклеотидные кодоны. Существует два вида наборов нуклеотидов. Триплеты, несущие информацию об аминокислотах — экзоны. Они образуют короткие сегменты ДНК и, как правило, разделены более длинными неинформативным участком нуклеотидов, которые называют интроны. В каждом гене число экзонов на один превышает число интронов. С началом транскрипции в состав и-РНК включаются как экзоны, так и интроны. Первичный транскрипт полностью копирует молекулу ДНК.
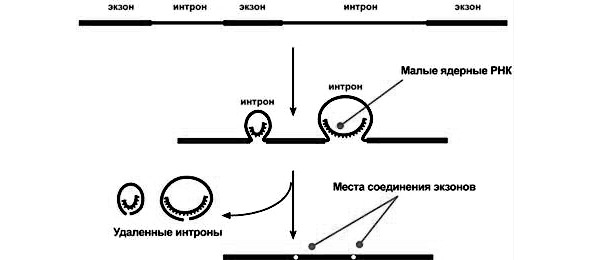
Транспортный уровень. Охватывает период между транскрипцией и трансляцией. На данном этапе происходит процессинг, то есть созревание и-РНК. Суть его — удаление интронов с помощью ферментов рестриктаз. Экзоны сохраняются и соединяются в единую цепь с помощью ферментов лигаз. Указанное явление называется сплайсинг. В ходе его возможны ошибки транскрипции, когда удаляются не все интроны. Прошедшая сплайсинг и-РНК значительно укорочена, компактна и может служить матрицей для синтеза белка. Для этого она переносится из ядра в цитоплазму к рибосомам. В клетках животных и растений на всем пути от ядерного хроматина к рибосомам и-РНК находится не в свободном состоянии, а связана специальными белками-переносчиками. Они различны в ядре и цитоплазме. Образно говоря, белки везут и-РНК в рибосому «на перекладных».
Трансляционный уровень. Трансляцией называется синтез полипептидной цепи из аминокислот согласно кодирующей и-РНК. В ходе трансляции происходит перевод генетической информации в аминокислотную последовательность: ДНК, и-РНК, белок. Трансляция является очень важной частью общего метаболизма клетки. В ней задействованы не менее 20 ферментов (аминоацилсинтетаз), до 60 различных т-РНК, 3—5 молекул р-РНК и макромолекулы и-РНК. Здесь выделяют следующие этапы: инициация, элонгация, терминация.
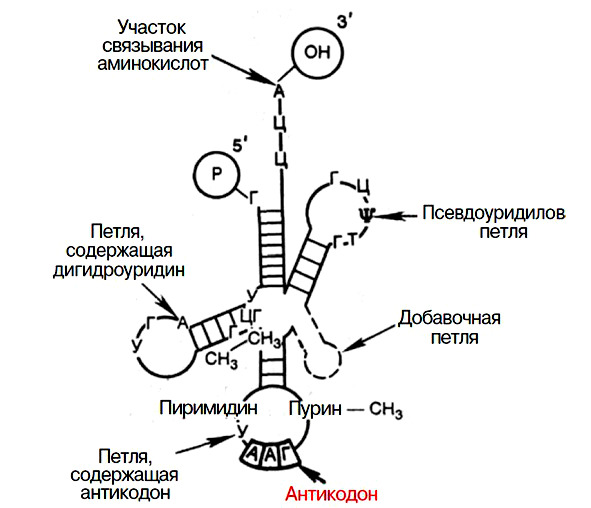
Инициация — начало трансляции. Фермент аминоацилсинтетаза активирует определенную аминокислоту и вовлекает ее в белковый синтез. Активированная аминокислота соединяется с т-РНК специализированной на данной аминокислоте. Эта т-РНК имеет форму трилистника и содержит три петли. Антикодоновая петля имеет антикодон, то есть триплет нуклеотидов, соответствующий своей аминокислоте. Псевдоуридиновая петля необходима при взаимодействии с рибосомой. Дигидроуридиновая петля служит для соединения с ферментом аминоацил-синтетазой. (Рис. 19). Доставленная к рибосоме и-РНК связывается с рибосомальной РНК. При этом рибосома диссоциирует, ее малая и большая субъединицы несколько раздвигаются, и рибосома насаживается на и-РНК. В начальной части и-РНК имеются стартовые кодоны АУГ и ГУГ. К ним присоединяется т-РНК, несущая соответствующую аминокислоту (рис. 20).
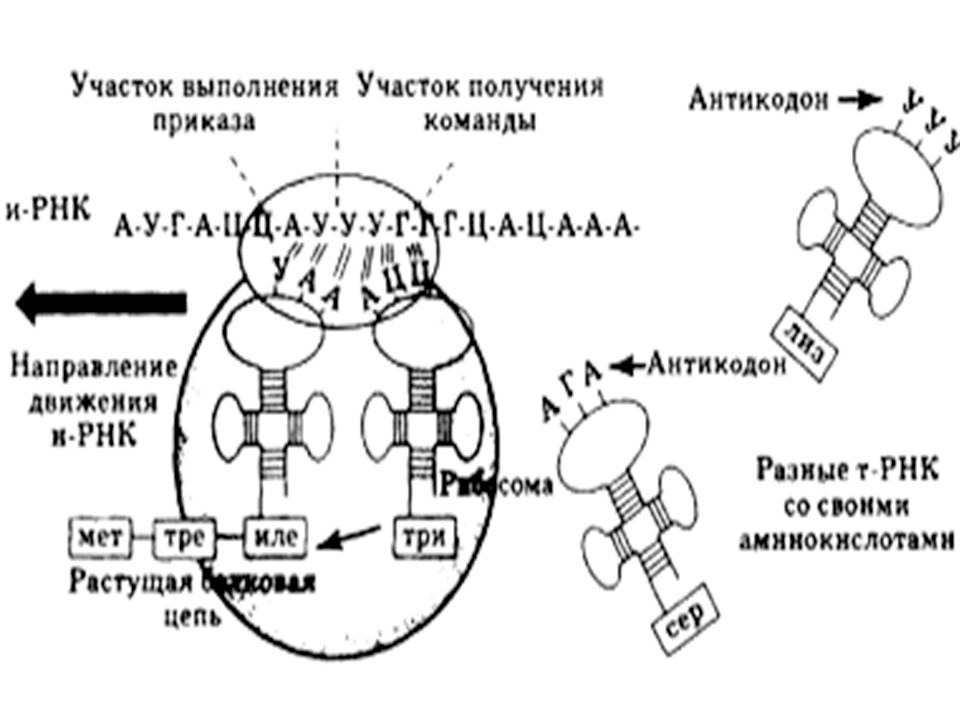
Элонгация — продолжение трансляции. Рибосома двигается вдоль молекулы и-РНК. Транспортные РНК связываются с рибосомой и их анти-кодоны избирательно по правилу комплементарности контактируют с кодонами и-РНК. При совмещении кодона и антикодона аминокислота отрывается от т-РНК и включается в полипептидную цепь из аминокислот в большой субъединице рибосомы. При формировании первичной белковой структуры каждая новая аминокислота занимает место за аминокислотой, включенной непосредственно перед ней.
Терминация — окончание синтеза. В и-РНК есть трейлерный участок, содержащий стоп кодоны УАА, УАГ, УГА. Они указывают на завершение синтеза данного белка. Поэтому при контакте с ними сборка аминокислотной цепи заканчивается. Большая и малая субъединицы рибосом смыкаются.
Посттрансляцнонный уровень. Синтезированная белковая молекула начинает усложнять свою первичную структуру под действием ферментов. Происходит ее конформация, изменяется пространственная организация белка, он приобретает вторичную, третичную и четвертичную структуру.
Таким образом, мы узнали центральную догму молекулярной биологии. Она гласит: информация в живых организмах передается по цепи ДНК — РНК — белок. В настоящее время доказано явление обратной транскрипции, когда передача информации происходит от РНК к ДНК. В то же время совершенно невозможен перенос информации от белков обратно к нуклеиновым кислотам.
В соответствии с существующими представлениями синтез белка всегда начинается с работы генов. Работа генов — это способность транскрибировать, то есть направлять синтез и-РНК. Но не всегда работа гена заканчивается сборкой белковой молекулы. Как заметил один американский генетик, для того, чтобы выдать белок, гену нужно пробиться сквозь «клеточные джунгли». Выход конечного белкового продукта — это экспрессия гена. Она совершается в результате деятельности всей клетки с ее многокомпонентными механизмами белкового синтеза.
Энергетический обмен
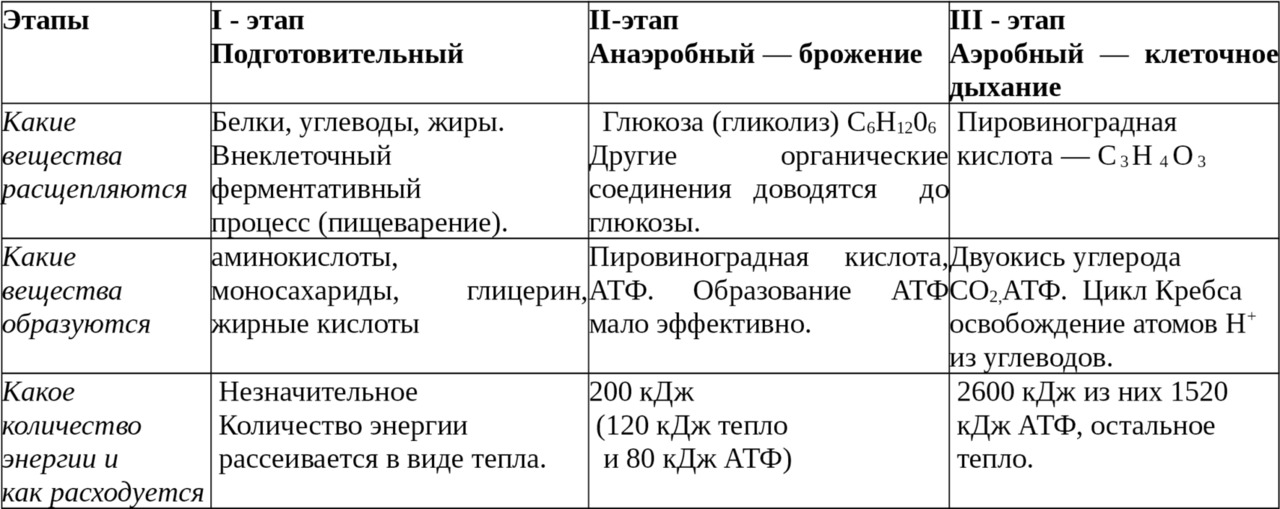
Энергетическим обменом или диссимиляцией называются процессы ферментативного расщепления органических веществ и образование соединений богатых энергией. Энергетический обмен подразделяется на три этапа.
Первый этап, подготовительный, связан с пищеварением. Он происходит вне клетки. Крупные молекулы биополимеров распадаются на мономеры: белки — на аминокислоты, полисахариды — на простые сахара, жиры — на жирные кислоты и глицерин. При разрыве химических связей выделяется небольшое количество энергии, рассеянной в виде тепла. Мономеры поступают в кровь.
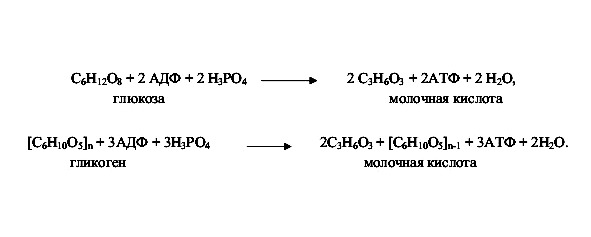
Второй этап — гликолиз, бескислородное расщепление глюкозы. Происходит внутриклеточно в цитоплазме, куда глюкоза поступает из крови. Включает ряд последовательных ферментативных реакций, в результате которых глюкоза распадается на две молекулы пировиноградной кислоты. Реакции протекают с участием фосфорной кислоты, образованием 2 молекул АТФ.
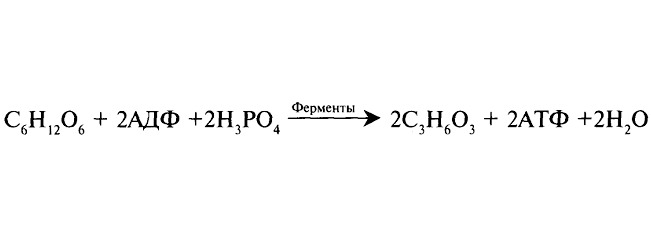
В процессе гликолиза выделяется 200 кДж энергии, из которых только 80 кДж (40%) аккумулируется в АТФ, остальные 120 кДж рассеиваются в виде тепла.
Гликолиз происходит во всех животных клетках, но является мало эффективным с энергетических позиций. Поэтому основные процессы накопления энергии совершаются на третьем этапе.
Третий этап — кислородный (аэробный — клеточное дыхание). Его называют окислительным фосфорилированием. Наблюдается полное кислородное расщепление органических веществ до двуокиси углерода СО2. Происходит освобождение атомов водорода Н (водород выделяется из углеводов в результате прохождения ими сложного ряда химических превращений, называемых циклом Кребса). Реакция протекает с участием АДФ и Н3Р04. При этом выделяется большое количество энергии, достаточное для синтеза 36 молекул АТФ.
Окислительное фосфорилирование совершается в митохондриях клеток Атомы водорода Н (электроны и протоны) переносятся на систему ферментов в митохондриальной мембране. Здесь они окисляются, то есть теряют электроны:
Н2 — 2е- 2Н+. Образуются свободные электроны е- и ионы водорода Н+ (протоны). В ходе дыхания электроны несколько раз пересекают мембрану митохондрий, вынося протоны Н+ на наружную поверхность. Количество положительно заряженных протонов там резко возрастает. Возникает градиент концентрации протонов и электрический потенциал. При напряжении 200 мВ в ферменте АТФ-синтетазе, встроенном в мембрану крист, открывается протонный канал. Через него протоны Н+ возвращаются на исходную позицию, где взаимодействуют с 02, образуя воду (2Н+ +02 = Н20). В момент прохождения протонов по каналу фермента электрическое поле разряжается, а энергия аккумулируется в реакции синтеза АТФ.
Итоговое уравнение внутриклеточного расщепления глюкозы:
Анаэробный этап:
Аэробный этап
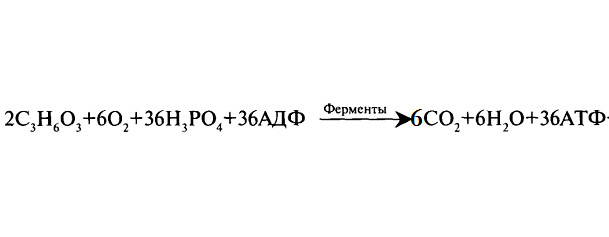
Суммарное уравнение гликолиза:
38АДФ +38Н3Р04 +1520 кДж = 38 АТФ +38Н20
Таким образом, в ходе энергетического обмена из одной молекулы глюкозы образуется 38 молекул АТФ.
Блок-схема энергетического обмена
Теория гена
Теория — это система обобщающих положений в той или иной области знаний. Теория неразрывно связана с практикой, которая ставит задачи и побуждает к их решению. Наука генетика наиболее выразительно иллюстрирует эту взаимосвязь. Успехи современной молекулярной биологии существенно углубили и детализировали теоретическую базу генетики, обосновали молекулярно-генетический уровень жизни.
Структура гена
Грегор Мендель, разрабатывая основопологающие законы наследования (1856—1863 гг.), использовал понятие «наследственные зачатки», применяя для них буквенные обозначения. Термин «ген» (от греч. genos — происхождение) ввел датский генетик В. Йогансен (1909 г.). Ген — структурная и функциональная единица наследственности.
В современном понимании ген — это участок молекулы ДНК (у некоторых вирусов — РНК) со строго определенной последовательностью нуклеотидов, контролирующей синтез белка (белков-ферментов).
Детальный анализ генной активности позволил выделить следующие группы генов:
1) аллельные и неаллельные;
2) доминантные и рецессивные;
3) эпистатические и гипостатические;
4) структурные и регуляторные.
В целом, понятие «ген» ассоциируется с созидательными процессами, однако есть и гены деструктивные по своей природе: онкогены, гены-мутаторы, летальные и сублетальные гены. Генам свойственно объединиться в группы, полигены. Функционально близкие группы генов формируют кластеры, отвечающие за важнейшие функции организма (размножение, пищеварение и т.д.).
На молекулярном уровне гены образованы нуклеосомами и связывающми их фрагментами молекулы ДНК. Нуклеосома состоит из протеинового дискообразного остова, включающего 8 молекул (глобул) белков-гистонов: по 2 молекулы H2A, H2B, H3, H4. На него насажен виток молекулы ДНК, включающей 150 пар нуклеотидов. Нить ДНК непрерывно и последовательно связывает нуклеосомы, при этом межнуклеосомные участки называются линкерными и каждый из них содержит до 60 пар азотистых оснований. Полная нуклеосома включает собственно нуклеосому и примыкающий к ней линкерный участок, насчитывая таким образом, около 200 пар нуклеотидов.
При спирализации ДНК свободные пространства заполняет белок Н1 (рис. 21).
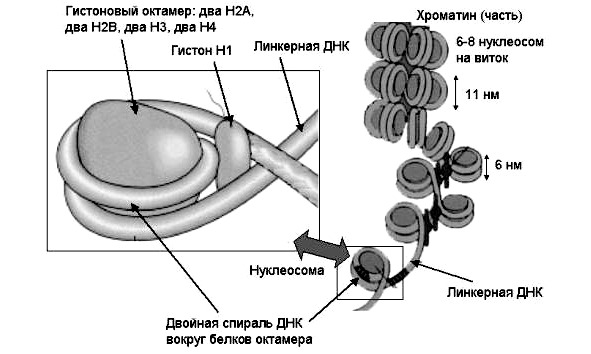
Ген средней величины объединят около 6 нуклеосом. Методами секвенирования было установлено, что организм человека содержит 25—40 тысяч активно работающих генов. В последнее время специалисты по биоинформатике уточняют количество генов до 2025 тысяч из-за их повторения в геноме. Следует учитывать высокую динамичность всей генной системы, и эти цифры, очевидно, будут меняться. Суть вопроса не в количестве генов, а в их сложности. Смысл эволюционных перестроек всей генной системы — это количество информации, включаемой в отдельный конкретный ген. Все гены функционируют как единое целое, формируя индивидуальный генотип особи и генотипическую среду, определяющую фенотипические проявления, т.е. признаки организма.
Генная регуляция синтеза белка. Система оперон
Основным условием существования любых живых организмов является наличие тонкой, гибкой и согласованно действующей системы регуляции, в которой все элементы тесно связаны друг с другом. В белковом синтезе определенное значение имеют не только количественный и качественный состав белков, но и параметры времени. Теорию генной регуляции синтеза белка разработали французские ученые Ф. Жакоб и Ж. Моно, удостоенные нобелевской премии (1965 г.). Для этого была использована культура бактерии кишечной палочки E. coli. Общая концепция состоит в способности этой бактерии включать или выключать систему генов в зависимости от наличия или отсутствия необходимой для жизнедеятельности бактерии лактозы (молочный сахар). Соответственно, система получила обозначение Lac-оперон (оперон — генетическая единица транскрипции). Она включает группу структурных и регуляторных генов.
Акцепторной зоной являются ген-промотор и ген-оператор. Структурные гены lac Z+, lac Y+, lac A+ содержат информацию о белках-ферментах, необходимых для расщепления лактозы: галактозидаза, пермеаза, трансацетилаза. Ген-регулятор регулирует образование регуляторного белка, контролирующего работу структурных генов. Ген-терминатор несет сигнал об окончании транскрипции (рис. 22).
Все указанные гены располагаются последовательно, за исключением гена-регулятора, которые занимают обособленное положение. Система работает рационально. В нерабочем состоянии ген-регулятор, контролирует выработку белка-репрессора (вещество-посредник), который находится в активной форме. Он включается в систему входа гена-промотора и далее связывается с геном-оператором, блокируя структурные гены. Механизм транскрипции закрыт. При поступлении в среду обитания Е. coli лактозы белок-репрессор переходит в неактивную форму, ген-оператор освобождается и структурные гены начинают механизм транскрипции. Происходит синтез ферментов расщепляющих лактозу как субстрат, необходимый для жизнедеятельности кишечной палочки. С полной утилизацией лактозы посредством активации белка-репрессора система посредством оператора опять блокируется. Таким образом, белок-репрессор является негативным регулятором.
Описанная для прокариот функциональная схема справедлива и для эукариотических клеток, хотя реализуется более сложными путями с участием гормонов. Кроме того, из-за наличия ядра в клетке процессы транскрипции и трансляции разделены не только пространственно биомембраной, но и во времени.
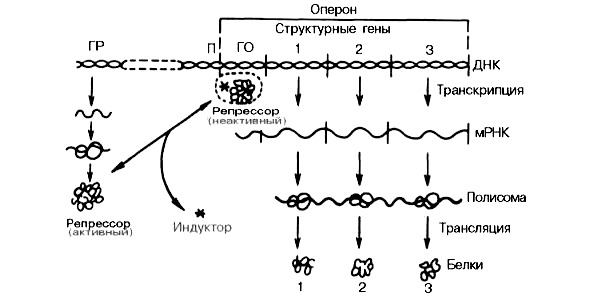
Хромосомы
Местом локализации генов в клетке являются хромосомы. Они относятся к числу самых удивительных внутриклеточных структур и с завидным упорством мигрируют из организма в организм на протяжении многих поколений. В миниатюрном биологическом компьютере, каким являются хромосомы, сконцентрирована информация в несколько терабайт, эквивалентная сотням томов обширных научных фолиантов. Здесь записано все — какими мы были, есть и будем.
Хромосомы — это высокоспециализированные компоненты клеточного ядра, обладающие особой индивидуальностью и функцией, способные к воспроизведению на протяжении ряда поколений. Свою четко выраженную морфологическую структуру хромосомы приобретают в ходе клеточного деления (митоза). Поэтому, все представленные ниже данные касаются митотических хромосом.
В состав хромосом входят ДНК, и-РНК, основные белки гистоны, негистоновые белки, Гистоны — это структурные белки относительно небольшого диаметра, несущие положительно заряженные аминокислоты. Положительный заряд способствует тесной связи гистонов с ДНК. Как указано выше, известно четыре типа гистонов, которые подразделяются на две группы: нуклеосомные гистоны Н2, Н3, Н4, и гистоны Н1 (см. рис. 21).
Строение хромосомы. В метафазе митоза хромосомы представлены палочковидными образованиями, сформированными подобно шпильке. В них различают плечи и центромеру, район первичной перетяжки. Расположение центромеры строго постоянно для определенной хромосомы.
В 1960 г. английский генетик Патау ввел понятие центромерный индекс — отношение длины плеча к длине всей хромосомы. В соответствие с этим выделены следующие виды хромосом:
1. Метацентрические хромосомы, имеющие срединно расположенную центромеру и плечи равной длины.
2. Субметацентрические хромосомы со смещенной центромерой и соответственно разновеликими плечами.
3. Акроцентрические хромосомы, у которых центромера резко сдвинута в крайнее положение, в связи с чем одно плечо почти редуцировано и определяется с трудом.
4. Телоцентрические хромосомы, где центромера замыкает хромосому с одного конца (рис. 23).
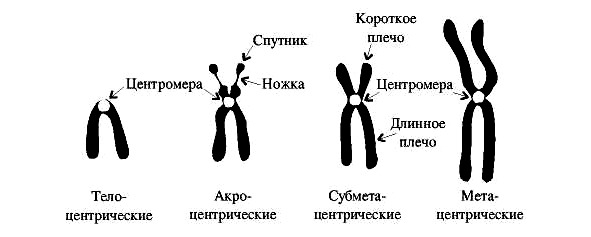
Появление на плече хромосомы вторичной перетяжки ведет к формированию дополнительного фрагмента плеча, называемого спутник.
Полностью сформированная хромосома состоит из двух морфологически одинаковых нитей — хроматид. Они переплетены, тесно прилежат одна к другой и соединяются с помощью центромеры. Свободные концы хромосом образованы теломерой, играющей роль своеобразного изолятора. Естественный конец хромосомы в виде теломеры не способен контактировать с другими хромосомами или их фрагментами. Теломера предотвращает отрыв и потерю нуклеотидных пар ДНК. Размеры метафазных хромосом у разных представителей животного мира колеблются в пределах 0,2—50,0 мкм. Длина хромосом человека 1,5—10,0 мкм. Количество хромосомных наборов в клетках является видовым признаком: у человека — 46, у кукурузы — 20, у мухи дрозофилы — 8 и т. д.
Общим принципом ультрамикроскопической организации хромосом является образование доменов (петель ДНК, связанных с белками). Они отходят под тем или иным углом от основной оси хромосомы. Типичная петля содержит от 20000 до 100000 пар нуклеотидов молекулы ДНК. Гигантская молекула ДНК, образующая совместно с белками хромосому, претерпевает сложную пространственную перестройку — «упаковывается», путем спирализации (см. рис. 24). В итоге общая длина ее уменьшается примерно в 10000 раз. С помощью белков каждая гигантская молекула ДНК компактно упаковывается и приобретает вид палочковидной структуры (рис. 24). Вся генетическая информация, заключенная в хромосоме, составляет геном. Каждая хромосома дифференцирована по длине и в ней выделяют два типа районов: эухроматиновые и гетерохроматиновые.
Эухроматин — это активные участки хромосом, содержащие основной комплексов генов. ДНК здесь пребывает в деспирализованном состоянии.
Гетерохроматин представляет собой неактивные в генетическом отношении фрагменты хромосом в резко конденсированной форме. Концевые теломеры всегда состоят из гетерохроматина.
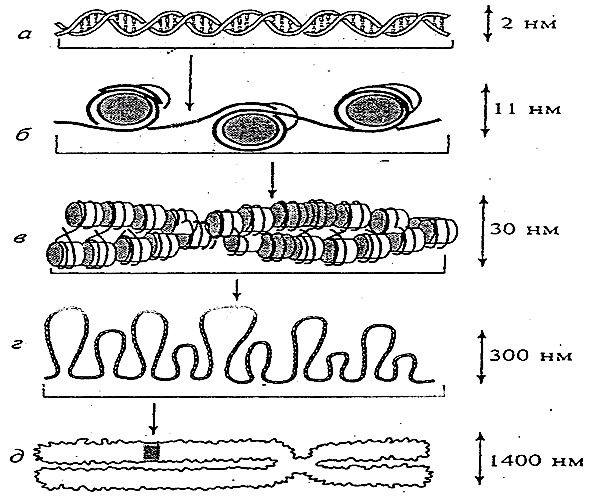
Интерфазные хромосомы находятся в деспирализованном состоянии и представляют длинные нити деконденсированной ДНК. В ядре клетки эти хромосомы всегда стремятся занять определенные области или зоны. Мнение, что они сплетены в единый клубок, согласно современным взглядом, маловероятно.
Многоклеточные организмы содержат два вида клеток: соматические (от слова soma — тело) и половые (генеративные) клетки — гаметы. Для ядер соматических клеток характерен двойной — диплоидный набор хромосом (2n). Зрелые половые клетки обладают половинным — гаплоидным набором хромосом (n), который составляет у человека 23 хромосомы. В диплоидном наборе хромосомы образуют идентичные пары. Две хромосомы в составе одной пары называются гомологичными. Они имеют одинаковые размеры, точно совпадающую локализацию центромеры и плечи идентичной формы и величины. У человека 46 хромосом и гомологичные хромосомы образуют 22 пары, называемые аутосомами. Последняя 23 пара представлена половыми хромосомами, определяющими половую принадлежность. У женщин эти хромосомы имеют форму XX, у мужчин XY и называются гетерохромосомами.
Хромосомы многоклеточных организмов подчиняются следующим правилам:
1. Все соматические клетки одной особи имеют в норме одинаковое число хромосом;
2. У всех организмов, относящихся к одному виду, число хромосом также одинаково. Хромосомный набор человека — 46, шимпанзе — 48, курицы — 78, рыбы сазан —104 хромосомы.
Кариотип и идиограмма
Совокупность количественных (число) и качественных (форма) показателей хромосом соматических клеток организма представляет его кариотип. Особи, относящиеся к одному виду, имеют одинаковый кариотип. Хотя у человека и шимпанзе число хромосом почти совпадает — это не является точным критерием их эволюционной близости. Одно и то же число хромосом может наблюдаться у очень далеких друг от друга организмов: муха домашняя, головная вошь, растение шпинат имеют в кариотипе по 12 хромосом. Число хромосом также не является показателем высоты организации (у курицы больше хромосом, чем у человека).
Если гомологичные хромосомы определенного кариотипа сгруппировать и расположить попарно в порядке убывания размеров получится кариограмма, а согласно положению центромеры — идиограмма. Идиограмма позволяет представить кариотип человека в виде схемы, которая включает семь групп, с буквенными обозначениями от А до G.

Чикагская конференция генетиков положила в основу классификации величину и расположение центромеры, а также длину плеч хромосомы: группа А — крупные метацентрические (1,2 и 3 пары), группа В — крупные субметацентрические (4 и 5 пары), группа С — средние субметацентрические (6—12 пары), группа D — крупные акроцентрические (13, 14, 15 пары), группа Е — маленькие субметацентрические (16, 17, 18 пары), группа F — маленькие метацентрические (19 и 20 пары), группа G — маленькие акроцентрические (21, 22 пары — Y).
В настоящее время существует множество технических приемов, которые позволяют детально разграничить хромосомы каждой пары и выявить даже незначительное отклонение в строении отдельных хромосом, а также локализацию в них генов.
Деление клетки
Клеточное деление — это удивительный биологический механизм, с помощью которого в ходе эволюции клетке удалось обмануть время и значительно продлить свое существование. Механизм деления клетки открыл любитель-натуралист лейпцигский продавец книг Вильгельм Гофмейстер в 1848 г. Он зарисовал отдельные стадии деления клеточного тела под микроскопом. Позднее Вильгельм Ру показал универсальность этого явления для животных и растений, что позволило выдающемуся представителю медицины Рудольфу Вирхову провозгласить: «Omnis cellula e cellulae» — каждая клетка из клетки
Деление клетки — митоз, представляет чрезвычайно сложное явление, посредством которого материал материнской клетки равномерно распределяется между дочерними клетками. То, что удается наблюдать при делении клетки в световом микроскопе, лишь внешнее отражение глубоких молекулярно-биохимических внутриклеточных процессов, протекающих в строгой последовательности. Эти процессы объединяет клеточный цикл. Он охватывает промежуток времени от одного деления клеток до другого деления с образованием двух новых клеток. Клетки различных типов имеют неодинаковую длительность клеточного цикла. Общая продолжительность его равна времени, в течение которого произойдет удвоение числа клеток определенной группы (клеточной популяции). Так, если исходное состояние популяции 2x10 клеток—10 часов, а удвоение 4x10 клеток—30 часов, то продолжительность клеточного цикла составляет 20 часов (30 час — 10 час = 20 час.)
В клеточном цикле выделяют: аутосинтетическую интерфазу и митоз (рис. 25).
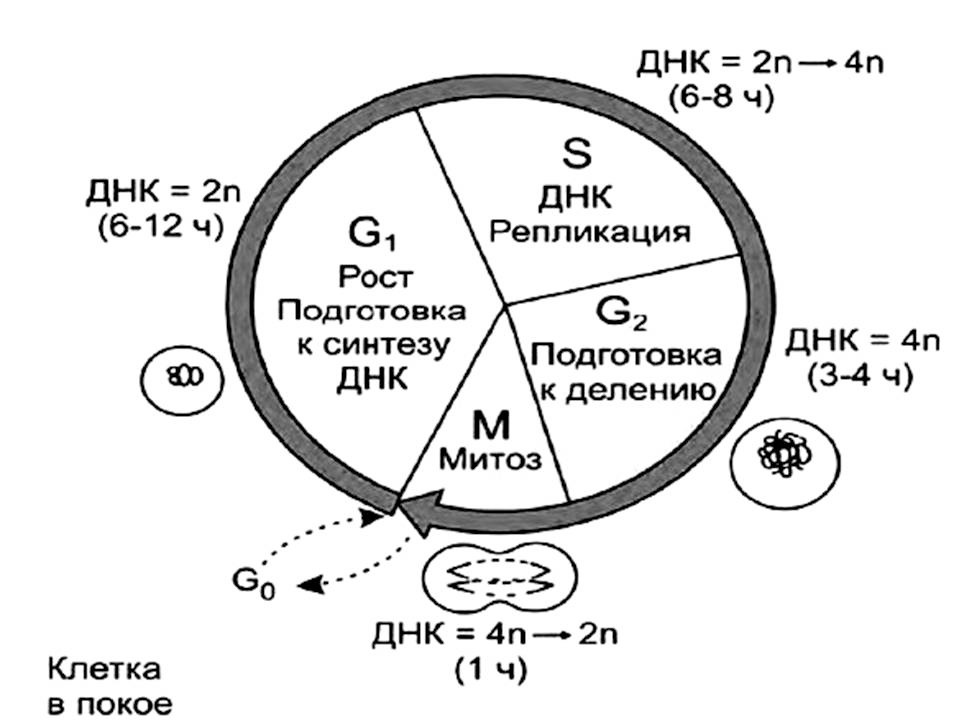
Аутосинтетическая интерфаза состоит из пресинтетического периода G1, синтетического периода S, постсинтетического периода G2.
В пресинтетическом периоде G1, клетка накапливает белки и РНК необходимые для формирования внутриклеточных структур. Здесь же может совершаться рост клетки. В G1 имеется точка старта (переходная точка). Это ключевой момент клеточного цикла, пройдя который, клетка включает механизм деления и должна осуществить все последующие стадии: S, G2, митоз. Подтверждением является простое наблюдение. Если клетку в S периоде, то есть прошедшую точку старта, совместить с другой клеткой в состоянии G1. (ранняя фаза), в последней немедленно начнутся процессы характерные для 8 периода. При дефиците основных питательных веществ клетки эукариот останавливаются в фазе G1 в точке старта. Продолжительность G1, максимальна и у разных клеток колеблется от нескольких часов до нескольких суток.
В постсинтетическом периоде G2 синтез ДНК прекращается, но наблюдается интенсивное образование АТФ и ядерных белков. Время необходимое для осуществления этих процессов 3—4 часа. Клетка начинает готовиться к сборке своего митотического аппарата.
Митотический аппарат клетки — это совокупность внутриклеточных структур, которые с началом митоза образуются из центросомы. Центросома (клеточный центр) — органелла, обеспечивающая течение митоза. Она состоит из двух центриолей, связанных тонкой перемычкой центродесмозой и окруженных лучистой сферой цитоплазмы — астросферой.
Митоз (М) — короткий период клеточного цикла. Сущность его заключается в конденсации содержимого ядра и выявлении в нем хромосом, которые удваиваются и затем равноценно распределяются между двумя дочерними клетками. Митоз — это непрерывный процесс, но для удобства рассмотрения в нем выделяют четыре фазы: профазу, метафазу, анафазу и телофазу (рис. 26).
Профаза. Начинается со сдвига физико-химического состояния цитоплазмы и реорганизации цитоскелета клетки. Центросома расщепляется. Ее центриоли расходятся в противоположные стороны, формируя два полюса клетки. Между ними образуется ахроматиновое веретено в виде тонких белковых нитей. Формируется мнтотический аппарат клетки, состоящий из микротрубочек и связанных с ними белков. Ядро клетки несколько увеличивается в размерах. В нем обнаруживаются хромосомы в виде тонких нитевидных структур. Вся группа хромосом выглядит в виде клубка. Ядрышки уменьшаются и исчезают. Конец профазы характеризуется распадом ядерной оболочки на мелкие фрагменты. К этому моменту хромосомы максимально укорачиваются и отделяются друг от друга.
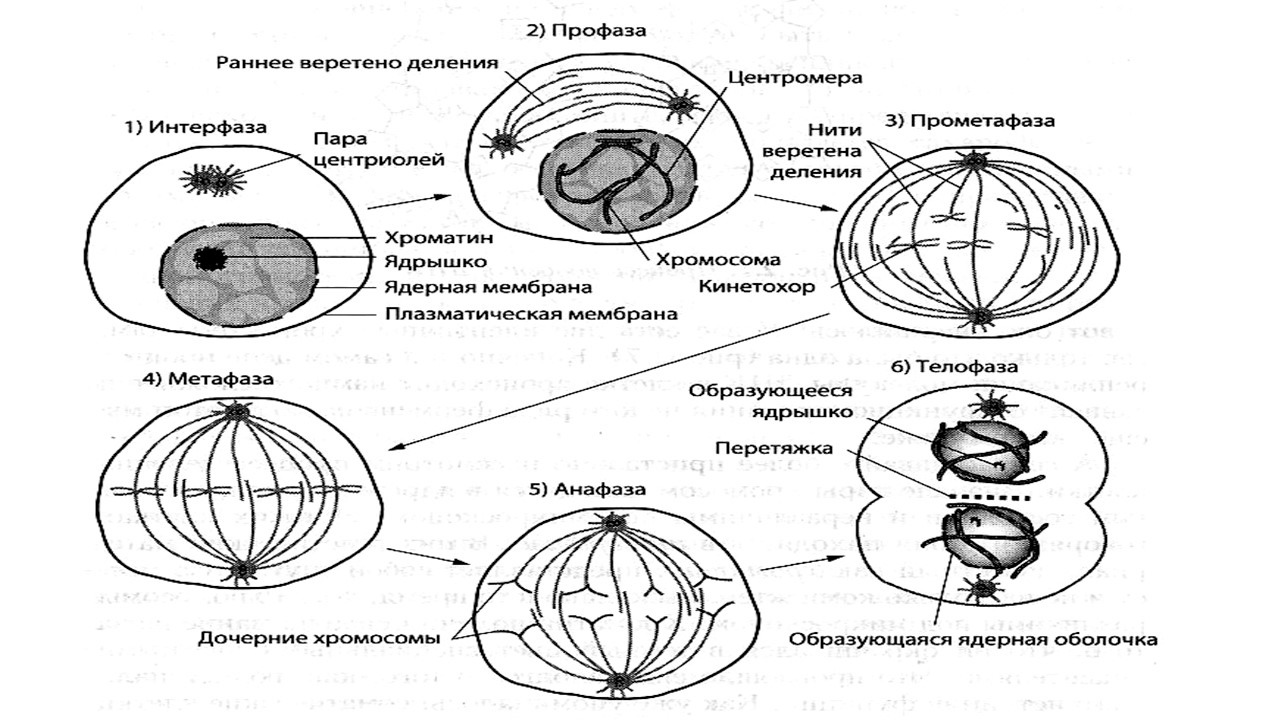
Метафаза. Хромосомы представлены компактными палочковидно изогнутыми тельцами. Каждая хромосома образована двумя тесно связанными половинками — хроматидами. Такие реплицированные (двойные) хромосомы прикрепляются к веретену с помощью особых структур, называемых кинетохоры. В метафазе хромосомы четко сгруппированы в центре клетки, их свободные концы обращены к периферии. Образуется фигура, называемая «материнской звездой» или метафазной пластинкой. Упорядоченное расположение хромосом в метафазной пластинке — это, главным образом, результат действия тянущей силы, создаваемой кинетохорными микротрубочками.
Анафаза. Началом ее является взаимное синхронное разделение всех хромосом на сестринские хроматиды, которые начинают движение к противоположным полюсам. Основное правило анафазы — хроматиды от одной хромосомы расходятся в разные стороны. Движение связано с укорочением микротрубочек кинетохора. В анафазе хроматиды называют дочерними хромосомами. Заканчивается анафаза сближением дочерних хромосом у противоположных полюсов, где они образуют две «дочерние звезды».
Телофаза — заключительная стадия митоза, связанная с реконструкцией ядер. Хромосомы деспирализуются и уже не различаются как отдельные морфологические структуры. Ахроматиновое веретено растворяется. Формируются путем сборки ядерные мембраны, начинается цитокинез — деление цитоплазмы. В животных клетках оно происходит путем гантелевидной перетяжки между ядрами, ее разрыва и образования двух дочерних клеток. Длительность всего клеточного цикла зависит от внешних и внутренних факторов, а также типа клеток. Наиболее короткой фазой является анафаза, максимальную продолжительность имеет интерфаза.
Регулирует и контролирует процессы митоза группа гормонов, относящаяся к факторам роста (см. ниже).
Отмечен суточный ритм митозов, связанных с биоритмами каждого организма. Так, у животных с ночным образом жизни пик митозов приходится на ранние утренние часы, у дневных животных и человека максимум митозов наблюдается в вечерние часы суток. Как долго исходная клетка может делиться митозом? Наблюдения над клетками соединительной ткани (нормальные фибробласты), выращенными в условиях лаборатории, показали, что способность каждой клетки данной линии делиться ограничена 50-ю удвоениями (лимит Хейфлика), после чего эта линия клеток погибает за счет исчезновения теломер. Если клеточную культуру заморозить на длительный срок после 10-го и 20-го удвоения, а затем разморозить, они все равно дадут только 50 удвоений. Также ведут себя и другие типы клеток, что обусловлено их генетической памятью. Клетки, как бы имеют «встроенные часы», точно отсчитывающие число делений своей линии. Жизнь клетки как любой живой системы ограничена.
В 1972 году ученый Ж. Керр впервые описал апоптоз — запрограммированную клеточную гибель. Апоптоз регулируется показателями внешней среды и рядом внутренних факторов, связанных с системой гомеостаза. Гомеостаз — постоянство внутренней среды организма. Посредством гомеостаза обеспечивается точный баланс различных видов клеток на определенных этапах онтогенеза.
Однако, из общего правила есть исключения. Культивируемую линию клеток можно превратить в «бессмертную», обработав вирусом рака. Сейчас существует около 600 бессмертных, трансформированных клеточных линий Наиболее известна линия клеток человека, называемая HeLa, которая возникла в 1952 г., в культуре ткани, взятой из матки женщины. С тех пор она непрерывно культивируется. Чтобы животные клетки приобрели способность к неограниченному клеточному делению, они должны обрести некоторые особенности раковых клеток.
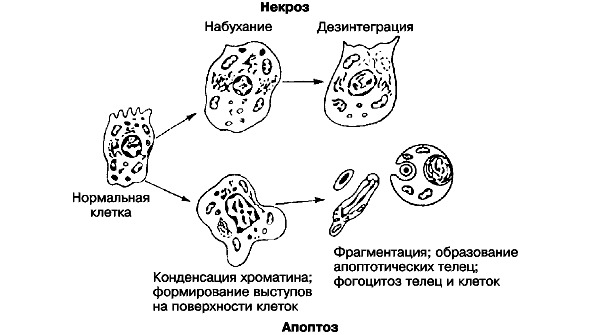
Биологическое значение митоза. Точный механизм клеточного деления обеспечивает качественное явление жизни — наследственность. В ходе митоза (клеточный цикл) происходит передача генетического материала в бесчисленных поколениях клеток. Митотическое деление клеток делает возможным рост и развитие. Оно поддерживает структурную целостность организмов посредством восстановления компонентов тканей, утраченных в ходе нормальной жизнедеятельности или при различных повреждениях.
Рост клетки
Закончив митоз, клетки вступают в рост. В процессе роста происходит увеличение объема ядра и цитоплазмы, формируется цитоскелет, закладываются клеточные органеллы. В итоге, каждая клетка приобретает свой ядерно-цитоплазматический коэффициент, характерный для данной клеточной линии:
Мn где: Мn — масса ядра,
Мс Мс — масса цитоплазмы.
Рост клеток контролируется факторов роста. Система включает: 1) белковый фактор роста, 2) специфические к нему клеточные рецепторы, 3) связывающие белки, которые регулируют поступление к клетке факторов роста. Помимо роста клеток, фактор роста оказывает свое действие на различные фазы клеточного цикла. Белковые факторы роста разнообразны и входят в группу гормонов роста. Особенностью их является то, что они вырабатываются не специальными железами, а неспецифическими клетками, находящимися во многих тканях. Наиболее изученным фактором роста является инсулиноподобный фактор. Он очень похож на белок инсулин, но ген инсулина находится у человека в хромосоме 19, а ген инсулиноподобного фактора роста — в хромосоме 15. Таким образом, эти белки являются продуктами различных генов.
Рост клетки сопровождается усилением пластического обмена и требует значительного напряжения энергетических процессов. На заключительном этапе роста клетка дифференцируется — приобретает специальные черты и становится функционально активной.
Раздел III. Организменный (онтогенетический) уровень организации живого/Размножение — фундаментальное свойство жизни
Размножением называется способность особи производить себе подобных, то есть давать потомство. Необходимость продолжения рода обеспечивается программой организма, которая имеет генетическую основу и выполняется при участии кластеров — интегрированных блоков генов. Это блоки эволюционной памяти, где отражены важнейшие этапы эволюции данного вида. Никто не учит живые организмы совершать порой чрезвычайно сложные действия для того, чтоб обеспечить появление и нормальное развитие своего потомства. Указанные особенности закреплены эволюционно и проявляются в виде инстинктов.
Составными элементами размножения являются наследственность, рост и развитие.
Наследственность обеспечивает тождество между родителями и потомками, тем самым, сохраняя непрерывность жизненного пути.
Рост — механизм, восстанавливающий дефинитивное состояние особей. В отсутствие роста любой способ размножения приводит к быстрому измельчению потомства и угасанию жизненного цикла в данной генетической линии.
Развитие — важный, но не обязательный элемент размножения. У некоторых примитивно организованных живых существ развитие выпадает, поскольку основные черты клеточной организации сохраняются.
Функция размножения отличаются от многих других функций тем, что служит интересам вида, а не индивидуума. В ходе эволюции жизненный успех сопутствовал тем видам, которые лучше размножались и давали более плодовитое потомство с лучшими шансами на выживание. Именно этим объясняется, почему многие особенности живых организмов, как структурные, так и поведенческие, подчинены интересам размножения, а в ходе онтогенеза для этого формируются специальные системы органов.
Типы размножения
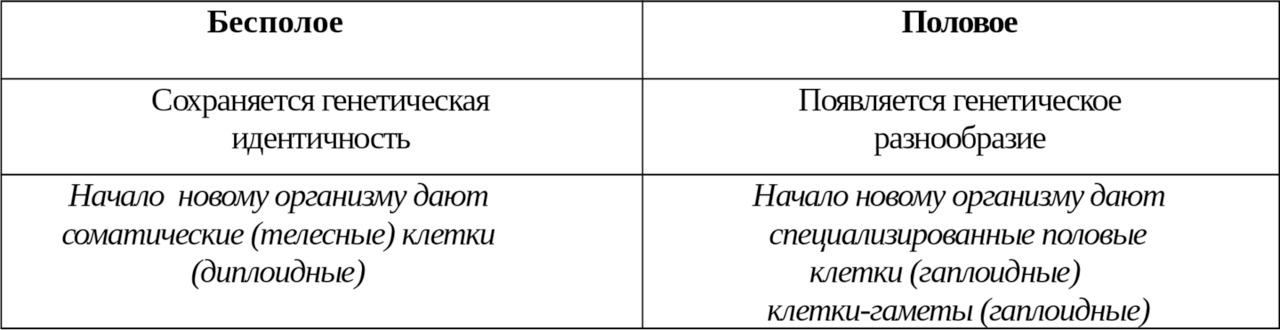
Бесполое размножение животных может происходить следующими способами.
1. Митотическим делением клетки. Это наиболее распространенный способ репродукции клеток, свойственный многоклеточным и одноклеточным организмам.
2. Простым делением клетки. Так размножаются, в частности, прокариотические клетки.
3. Шизогонией, или множественным делением, которое представляет более редкую форму, свойственную, например, некоторым простейшим организмам.
4. Почкованием клетки, которое бывает наружным и внутренним. При наружном почка образуется на поверхности клетки, при внутреннем изнутри, причем перед отделением почки поступают в специальную полость внутри материнской клетки и лишь затем выводятся наружу. Данный способ хорошо выражен у сосущих инфузорий — сукторий.

5. Спорообразованием, репродукцией при помощи спор, форма типичная для простейших споровиков.
6. Вегетативное размножение характеризуется тем, что начало новому организму дают группы клеток или многоклеточные зачатки, иногда участки тела родителя. Вегетативный способ наиболее широко распространен у низших представителей животного царства (кишечнополостных, губок), но может наблюдаться и у относительно высокоорганизованных животных (кольчатых червей).
Половое размножение животных получило в природе доминирующие положение, потому что является неиссякаемым источником изменчивости в результате перекомбинаций генетического материала. Оно совершается с участием половых клеток, гамет.
Половые клетки, гаметы, являются высокоспециализированными клетками, предназначенными для полового размножения. Их главная функция — осуществить перенос в закодированном виде генетической информации от родительских форм к потомкам, обеспечить формирование зиготы. Тем самым, с помощью половых клеток реализуется механизм наследственности.
Известны две группы половых клеток: мужские гаметы — сперматозоиды (спермии) и женские гаметы — яйцеклетки (овоциты, яйцевые клетки). Первые способны к различным по форме и быстроте типам движений, вторые — неподвижны, имеют систему оболочек и могут включать различные количества питательного материала необходимого для развития зародыша.
Принципиальные отличия половых клеток от соматических (телесных) состоят в следующем:
1. Зрелые гаметы гаплоидны, то есть содержат половинный набор хромосом.
2. В половых клетках резко изменены объемные соотношения ядра и цитоплазмы (ядерно-цитоплазматический коэффициент). В закончивших развитие половых клетках процессы метаболизма замедлены.
Развитие половых клеток
Развитие половых клеток, гаметогенез, включает процессы сперматогенеза и овогенеза. Эти процессы происходят в половых железах, гонадах, которые называются соответственно семенники и яичники.
Сперматогенез
Развитие мужских половых клеток включает четыре стадии: размножение, рост, созревание, формирование.
У человека все они совершаются в семенниках и продолжаются с периода полового созревания до старости (рис. 28).
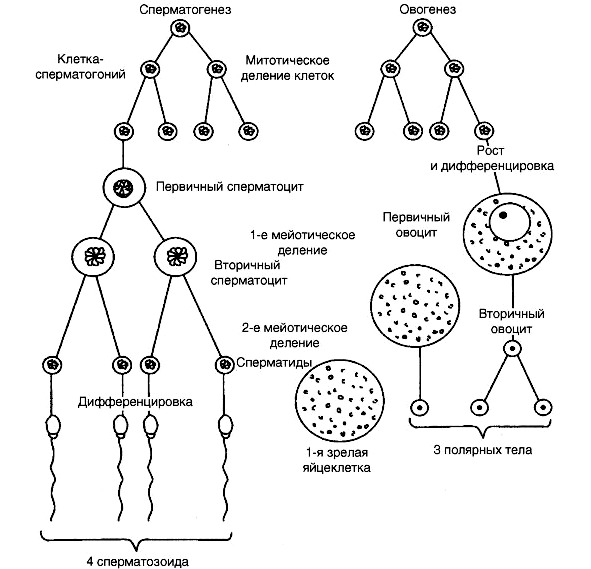
Размножение. В период полового созревания у лиц мужского пола в канальцах семенников начинается митотическое деление диплоидных первичных половых клеток сперматогоний. Этот процесс продолжается непрерывно десятки лет. В результате количество клеток существенно возрастает.
Рост. На стадии роста размеры клеток несколько возрастают, и они превращаются в сперматоциты. Важным событием этого этапа является репликация ДНК при сохранении неизменным числа хромосом.
Созревание. Главным для данной стадии служит процесс мейоза, включающий два последовательных клеточных деления с образованием четырех гаплоидных клеток сперматид.
Формирование. Путем сложных преобразований у сперматид формируется жгутик — специальный органоид движения и образуются зрелые половые клетки спермии (сперматозоиды).
Овогенез
Развитие женских половых клеток происходит в яичниках женщины и заканчивается в маточных трубах. Здесь выделяются три стадии: размножение, рост, созревание.
В организме женщины овогенез начинается уже в период эмбрионального развития, но затем приостанавливается до начала полового созревания.
Размножение. Суть процесса — деление путем митоза диплоидных первичных половых клеток — овогоний. В результате в яичниках закладывается значительный резерв клеток для последующего развития.
Рост. Процессы роста начинаются с периода полового созревания и имеют целью создать в половой клетке запасы питательных веществ. В результате величина клеток значительно возрастает, и они превращаются в овоциты. На этом этапе происходит также репликация ДНК при сохранении неизменным числа хромосом. В яичниках человека процессы роста сопряжены с увеличением фолликулов, в которых находятся половые клетки (рис. 29).
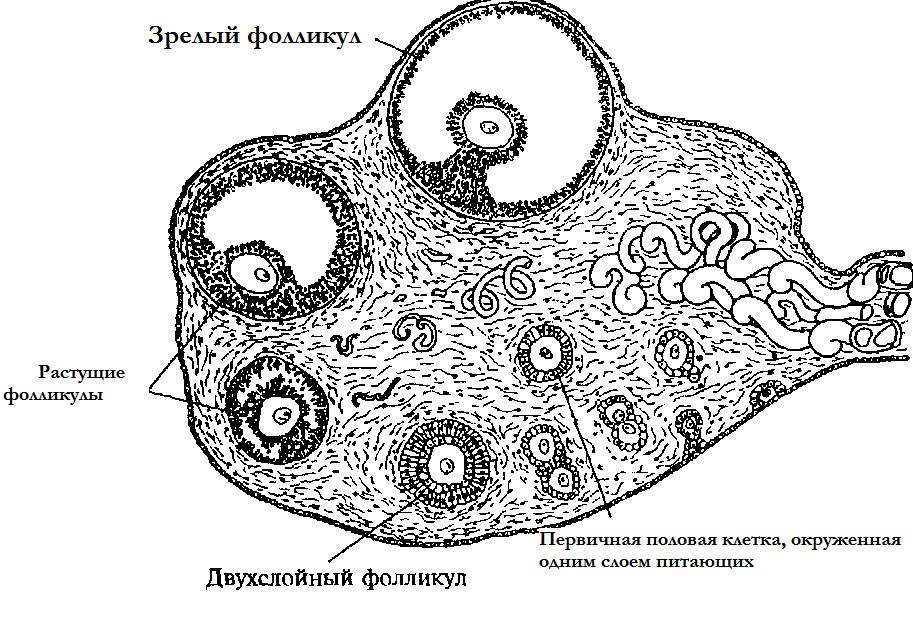
Созревание. На данной стадии совершается мейоз, существенно отличающийся от мейоза в семенниках. Первое деление мейоза заканчивается образованием двух гаплоидных клеток: крупного овоцита и мелкого первичного полоцита. Второе деление мейоза (происходит у млекопитающих животных и человека в ходе оплодотворения) сопровождается образованием крупной зрелой яйцеклетки и вторичного мелкого полоцита. Таким образом, в процессе мейоза образуется только одна полноценная женская гамета, где концентрируется весь питательный материал — желток. Три полоцита в размножении не участвуют.
Мейоз
Это центральное событие гаметогенеза, в результате которого незрелые диплоидные половые клетки с набором хромосом — 2n, превращаются в зрелые гаметы с набором хромосом — n.
Мейоз включает два последовательных клеточных деления. Первое называют редукционным, так как оно приводит к возникновению гаплоидных клеток. Второе деление — эквационное (уравнительное), в результате чего количество гаплоидных клеток увеличивается.
Синтез ДНК предшествует только первому редукционному делению. В ходе деления мейозом наблюдаются такие же фазы, как при митозе: профаза, метафаза, анафаза, телофаза.

Первое деление мейоза (редукционное)
Профаза. Происходит формирование веретена деления клетки. Процесс спирализации хромосом протекает сложно и имеет ряд особенностей, в связи с чем здесь выделяется несколько стадий:
Лептотена — увеличение ядра клегки, начало спирализации хромосом, которые видны в виде тонких нитей в нуклеоплазме (а).
Зиготена — хромосомы хорошо контурированы, гомологичные хромосомы сближаются (конъюгируют) и образуют пары — биваленты (б).
Пахитена — каждая хромосома в биваленте подразделяется на две четко выраженные хроматиды, в результате чего формируются тетрады (греч. tetra — четыре). На этой стадии происходит кроссинговер — обмен участками хроматид гомологичных хромосом. Обмениваться фрагментами могут как внутренние, так и внешние хроматиды (в).
Диплотена — гомологичные хромосомы расходятся в области центромер. Но связь в точках перекреста (хиазмы) сохраняется (г).
Диакинез — ядерная оболочка исчезает, а гомологичные хромосомы, отойдя друг от друга, удерживаются лишь в точках хиазм и приобретают форму колец, петель или крестов (д).
Метафаза. Формирование веретена деления клеток завершено. Хромосомы прикреплены к нитям веретена центромерами и располагаются по экватору клетки (е).
Анафаза. Силы притяжения между гомологичными хромосомами падают, и происходит синхронное расхождение гомологичных хромосом к противоположным полюсам веретена деления клетки. При этом в разные стороны отходят целые гаплоидные наборы хромосом. Каждая хромосома имеет две хроматиды (ж).
Телофаза. У полюсов клетки формируется по гаплоидному (одинарному) набору хромосом, но каждая из хромосом содержит двойное количество ДНК, поскольку состоит из двух хроматид (з). Короткий интеркинез (и).
Формула генетического материала будущих дочерних клеток — n2с.
Второе деление мейоза (эквационное)
Второе деление обычно начинается сразу после первого деления мейоза. Синтез ДНК перед этим делением не происходит. Характер деления напоминает митоз. Однако, особенностью его является то, что клетки, вступающие в деление, обладают гаплоидным набором хромосом.
Профаза и метафаза совершаются обычным путем. Анафаза имеет принципиальное отличие от первого мейотического деления. К полюсам веретена клетки расходятся хроматиды от одной хромосомы (в анафазе первого деления расходились целые хромосомы из гомологичных бивалентов). В итоге второго деления телофазные дочерние клетки сохраняют гаплоидный набор хромосом, но количество ДНК уменьшается в них в два раза (рис. 30, к, л, м).
Таким образом, в результате мейоза из одной исходной клетки образуется четыре половые клетки. Их итоговая генетическая формула nс.
Обратим внимание на дальнейшее преобразование генетического материала в половых клетках при оплодотворении.
В проникшем в яйцеклетку сперматозоиде при образовании мужского пронуклеуса происходит синтез ДНК. Синхронно с этим происходит синтез ДНК и в женском пронуклеусе. Зрелые пронуклеусы имеют генетическую формулу — n2с. при их слиянии образуется диплоидная зигота 2n4с. Зигота делится путем митоза на две клетки (бластомеры), с формулой 2n2с. Так завершаются сложнейшие преобразования материала наследственности.
Биологическое значение мейоза
1. В результате мейоза из диплоидных первичных половых клеток образуются гаплоидные зрелые половые клетки — гаметы.
2. В ходе мейоза происходит перекомбинация генетического материала в половых клетках, что обеспечивает наследственную изменчивость организмов.
3. Мейоз приводит к образованию сперматозоидов с разными половыми хромосомами X, Y, что в дальнейшем является решающим при формировании пола в процессе оплодотворения.
Строение половых клеток
Яйцеклетки имеют округлую форму, систему оболочек, ядро с гаплоидным набором хромосом (пронуклеус) и овоплазму, в которой может содержаться определенное количество включений, необходимых для развития зародыша. В зависимости от количества желточных включений размеры их варьируют от мкм до см. Типы яйцеклеток:
1. Изолецитальные яйцеклетки, мелкие, с очень небольшим количеством желточных включений относительно равномерно распределенных в цитоплазме. Ядро обычно занимает центральное положение в клетке (яйцеклетки моллюсков, иглокожих, ланцетников и млекопитающих).
2. Телолецитальные яйцеклетки, более крупные, богатые желтком. Включения распределены неравномерно, большая часть их концентрируется в одной из полусфер, где образуется вегетативный полюс клетки. Ядро находится в противоположной, бедной желтком зоне — анимальный полюс. Различают умеренно телолецитальные (у амфибии) и резко телолецитальные (у птиц) яйцеклетки.
3. Центролецитальные яйцеклетки, в них ядро занимает центральное положение, а вокруг располагаются желточные зерна. Поверхностные слои цитоплазмы свободны от желтка и перед оплодотворением сюда мигрирует ядро (яйцеклетки членистоногих).
В процессе исторического приспособления животных к условиям существования происходило усложнение организации яйцеклеток. Изменялось содержание желтка, имело место перестройка яйцевых оболочек. Различают первичные, вторичные и третичные оболочки яйцевых клеток. Первичная оболочка (желточная) вырабатывается самой яйцеклеткой и представлена тонкой цитоплазматической мембраной, придающей яйцеклетке определенную форму. Вторичная оболочка — продукт деятельности фолликулярных (питающих) клеток яичника, окружающих овоцит. Она хорошо выражена у яйцеклеток человека, млекопитающих животных и называется прозрачной оболочкой. Третичная оболочка формируется за счет секретов полового тракта при перемещении яйцевой клетки по половым путям (студенистая оболочка яиц амфибий, скорлуповая оболочка яиц птиц).
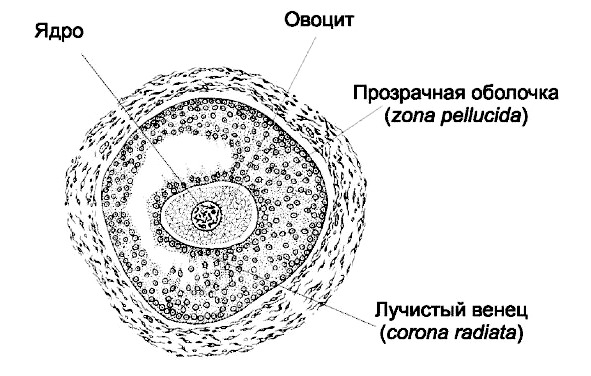
Сперматозоиды — мелкие подвижные клетки, длина которых измеряется в микрометрах. В сперматозоидах человека выделяют ряд отделов: головку, шейку, тело и жгутик (хвост), который заканчивается концевой нитью. Снаружи клетка покрыта плазмалеммой. Головка уплощена, имеет размеры 6х4 мкм, содержит ядро с нечеткой ядерной оболочкой и особым образом конденсированным хроматином. ДНК имеет кольцевую форм.
Начальный участок головки прикрыт акросомой в виде уплощенного мешочка. В нем находится акросомный матрикс богатый гидролитическими ферментами (гиалуронидаза, протеаза). В короткой шейке содержится центросома, от которой начинается аксонема жгутика. Тело длиной 6—10 мкм богато митохондриями, расположенными по спирали вокруг осевой нити аксонемы. Митохондрии связаны со специфическим белком убиквитином, который служит маркером отцовских митохондрий и способствует их разрушению после формирования зиготы. Жгутик состоит из аксонемы и добавочных парааксонемных структур. Длина его 50 мкм. В основе аксонемы лежит система микротрубочек (центральная пара и 9 периферических пар-дуплетов). Движение жгутика происходит за счет скольжения соседних дуплетов относительно друг друга (рис. 32).
При изучении микропрепарата у человека можно обнаружить наряду с нормальными определенный процент гигантских и карликовых форм, а также конические, грушевидные и двухголовые сперматозоиды. Состав популяции мужских половых клеток у отдельных индивидуумов остается стабильным в течение длительных периодов жизни. Эякулят фертильных лиц должен содержать не менее 30% нормальных сперматозоидов.
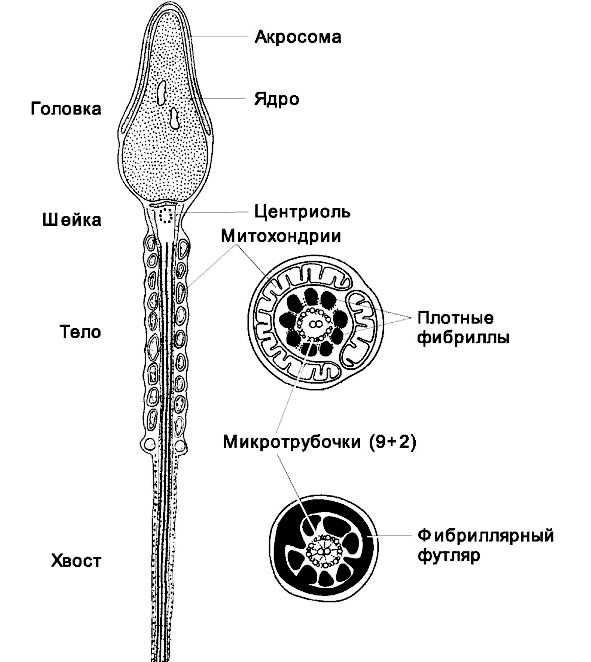
Онтогенез и его периодизация
Онтогенез — это индивидуальное развитие организма от момента формирования зиготы до естественного окончания жизни.
В основе онтогенеза лежит последовательная передача наследственной информации на всех этапах существования организма в определенных условиях среды. Онтогенез различных представителей животного мира отличается общей продолжительностью, скоростью и характером дифференцировок и детерминаций. Существуют различные схемы периодизации онтогенеза, применяемые для решения конкретных научных задач. Одной из наиболее распространенных является эколого-эмбриологическая схематизация. Согласно ей, онтогенез подразделяется на два основных периода — эмбриональный и постэмбриональный.
Первый период — эмбриогенез охватывает промежуток времени от оплодотворения до завершения основных процессов формирования тканей и органов. В зависимости от типа онтогенеза конец эмбрионального периода связывают с рождением (внутриутробный тип), выходом из яйцевых оболочек (личиночный тип) или освобождением от зародышевых оболочек (неличиночный тип развития).
Второй период — постэмбриональный следует за эмбриональным и отражает дефинитивное состояние организма.
Эмбриогенезу предшествует прогенез, или предзародышевый период, когда происходит развитие половых клеток.
Применительно к человеку употребляют термины — антенатальный (дородовый) и постнатальный (послеродовый) периоды жизни.
Эмбриогенез протекает стадийно. В нем выделяют следующие этапы: 1) оплодотворение, 2) дробление, 3) гаструляция — образование зародышевых листков, 4) гисто- и органогенез, формирование тканей и органов.
Эмбриональное развитие. Оплодотворение
Оплодотворение — это слияние сперматозоида и яйцеклетки, в результате чего возникает зигота, новый организм с диплоидным набором хромосом.
Оплодотворению предшествует осеменение, совокупность процессов, ведущих к встрече мужских и женских гамет. В естественных условиях осеменение бывает наружным или внутренним. При наружном яйцеклетки и спермии выводятся из организма, во внешнюю среду, при внутреннем, спермии вводятся в половые пути самки путем спаривания животных.
У представителей разных систематических групп животного царства оплодотворение имеет свои особенности, что связано с экологией, характером осеменения, типом яйцевой клетки, морфологией и физиологией половой системы самцов и самок. Несмотря на это, можно выделить общие биологические закономерности данного процесса у животных. Оплодотворение не мгновенный акт, его подразделяют на три последовательные фазы.
I фаза — сближение гамет в пространстве. Своевременный контакт спермиев и яйцеклеток имеет исключительно важное значение для нормального оплодотворения, так как зрелые гаметы, выделенные из половых желез, быстро «стареют» и через определенное время теряют оплодотворяющую способность. Встрече половых клеток способствуют следующие факторы: 1) биологически активные вещества гомоны, продуцируемые в окружающую среду зрелыми половыми клетками (фертилизин), 2) движение спермиев, характер которого может быть поступательно-прямолинейным при внутреннем осеменении или спиральным в случае наружного осеменения, 3) разность биоэлектрических потенциалов яйцевой клетки и среды, включающей мужские половые клетки. Вероятность встречи гамет зависит от числа спермиев, выделяемых при половом акте самцом. Многие виды животных продуцируют избыточное количество мужских половых клеток.
II фаза — контакт гамет — сингамия, проникновение спермия в яйцеклетку и ее ответная реакция. Несмотря на то, что в оплодотворении участвует большое число спермиев, непосредственно в яйцеклетку в норме проникает обычно один (моноспермия). Лишь у некоторых видов животных в яйцо попадает несколько спермиев (полиспермия) У разных видов это происходит на определенных стадиях созревания овоцита.
При взаимодействии мужской и женской гамет возникает акросомная реакция. Суть ее состоит в морфофункциональных изменениях акросомы спермия, в результате чего выделяются акросомальные ферменты (гиалуронидаза, трипсиноподобный энзим и др.), необходимые для оплодотворения. У многих животных имеет место выталкивание из акросомы более или менее длинной нити, которая попадает в особый воспринимающий холмик на поверхности яйцеклетки. Акросомная реакция способствует не только внедрению спермия, но и служит предпосылкой для реализации процессов, активирующих яйцо, то есть побуждающих его к развитию.
Контакт спермия с мембранами яйцевой клетки сопровождается кортикальной реакцией активации яйца. Суть ее в распаде или уменьшении плотности мелких кортикальных гранул на поверхностном слое овоплазмы. Эти гранулы имеются у представителей многих групп животных, в том числе далеких в систематическом отношении. Импульс активации приводит также к изменению биоэлектрической активности, связанной с физико-химическими процессами в поверхностных слоях овоплазмы. Видимым отражением этого у некоторых животных служит образование особой оболочки оплодотворения. Она является надежным механизмом, блокирующим проникновение в яйцо дополнительных спермиев при моноспермном оплодотворении.
III фаза — формирование мужского и женского пронуклеусов и образование зиготы. Ядерное вещество спермия (мужской пронуклеус) в овоплазме быстро увеличивается в размерах и достигает примерной величины женского пронуклеуса. Они сближаются и сливаются, в результате чего возникает синкарион — ядро зиготы.
Зигота — качественно новая клетка с диплоидным набором хромосом. В ней резко активированы процессы жизнедеятельности (степень метаболизма, уровень тканевого дыхания и др.), связанные с основной функцией нового организма — сохранением индивида и вида.
Особенности оплодотворения у человека и млекопитающих животных
Оплодотворение у человека происходит в маточных трубах, у млекопитающих животных в яйцеводах. Указанные органы женской половой системы обеспечивают продвижение яйцеклеток и ранних зародышей из яичников по направлению к матке (рис. 33).
У животных встреча мужских и женских гамет строго синхронизирована во времени и в пространстве. Это связано с наличием у самок млекопитающих ритмичных половых эстральных циклов, развитием в определенной стадии цикла половой доминанты и физиологическими механизмами осеменения. Спаривание животных возможно в конкретной фазе цикла (эструс), видимым проявлением которой является течка (половая охота). Введенные в половые пути спермии быстро достигают яйцеводов. К этому моменту закончен рост фолликулов яичников — пузырьковидных образований, содержащих овоциты. Происходит разрыв фолликулов (овуляция), и овоциты попадают в яйцеводы (у некоторых видов овуляция возникает в результате спаривания животных). В силу названных особенностей осеменения контакт гамет — сингамия всегда происходит у животных только в яйцеводах.
У человека эволюция репродуктивной функции привела к возникновению полового менструального цикла. Воспроизведение подчинено воле, половой процесс контролируется сознанием и не зависит от стадии цикла. Поэтому поступление спермиев в маточные трубы имеет произвольный характер и не всегда совпадает с выходом овоцитов из фолликулов яичников. Контакт может произойти не только в любом отрезке маточных труб, но и матке. Соответственно возрастает вероятность участия в оплодотворении стареющих гамет, мужских или женских. Это значительно усиливает опасность нарушения эмбриогенеза.
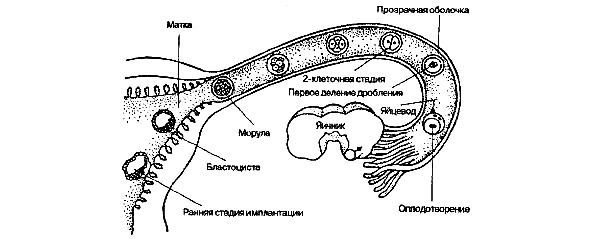
Для млекопитающих животных и человека характерен феномен капацитации спермиев. Капацитация предшествует внедрению мужской клетки в яйцо и представляет начальную стадию активации спермия. С его поверхности удаляются белки семенной жидкости, затем изменяются гликопротеиды плазматической мембраны. Заключительная фаза активации — акросомная реакция, при которой ферменты акросомы — гиалуронидаза, акрозин и другие выделяются в окружающую среду. Под влиянием указанных ферментов в местах контакта спермия с прозрачной оболочкой (вторичная оболочка) яйца происходит расщепление гиалуроновой кислоты и некоторых белков, что облегчает прохождение спермия в околожелточное пространство. Здесь спермий соединяется с плазматической мембраной овоплазмы и прикрепляется к ней постакросомальной областью. Это наиболее активный фрагмент мужской половой клетки, так как именно он формирует смешанный (аллогенный) участок плазматической мембраны мужской и женской гамет.
Спермий оказывается включенным в состав овоплазмы, что сопровождается реакцией активации яйца (рис. 34). Суть ее — образование новой плазматической мембраны из мембран распавшихся кортикальных гранул. Указанная мембрана блокирует полиспермию (у многих видов млекопитающих блок полиспермии осуществляется еще на стадии прохождения спермия через прозрачную оболочку).
Бесплатный фрагмент закончился.
Купите книгу, чтобы продолжить чтение.