
Бесплатный фрагмент - Второй вызов официозу: Палеоантропология
Все права защищены. Никакая часть электронного экземпляра этой книги не может быть воспроизведена в какой бы то ни было форме и какими бы то ни было средствами, включая размещение в сети Интернет и в корпоративных сетях, для частного и публичного использования без письменного разрешения владельца авторских прав. Автор имеет право вносить изменения и дополнения.
Вступление
Данная книга — о «скользких» вопросах в данной науке, о вопросах, положенных «под сукно», о научных заблуждениях и последних открытиях. Факты, факты и ничего, кроме фактов!
Например о том, как признали ископаемый окаменелый мозг геологической конкрецией, или как нашли другой окаменелый мозг, чего-де в природе «быть не может», о рогатых представителях рода человеческого, и о древних гигантах, не признаваемых официальной наукой, или о набившем оскомину «снежном человеке» (троглодите), а также о малоизвестном виде человекообразной обезьяны.
Каждый читатель вправе сам делать собственные заключения.
От автора
Глава 1 Сверхчеловек. Сегодня ли это уже?
Порой в наем довольно рафинированном обществе заходит речь о сверх-нормальных способностях человека. Во многом необъяснимая пока экстрасенсорика тому пример. Однако, наука и здесь делает первые шаги в попытке объяснить таинственные причины, позволяющие производить оздоровляющее воздействие посредством обычного взгляда или манипуляций рук. Так, например, своё объяснение поучил факт т. н. «заряжания» энергией воды и пр. жидкостей и воздействия её на организм: экстрасенс концентрирует в себе магнитную энергию, расщепляющую крупные молекулы, вследствие чего жидкость становится более «активной», что благотворно воздействует на живой организм.
Но задумывался и кто когда-нибудь, чем же всё-таки вызвано появление людей с неординарными возможностями, вроде бы и нехарактерными для человека? Возможно здесь следует искать ответ не только и не столько в природе человека, а в тех биологических процессах эволюции, которые, как известно, неспешно, поэтапно, но кардинально меняют мир. Вспомним, прошли эпохи моллюсков, земноводных, динозавров… Так, может быть, появление сверх-способностей есть необходимая закономерность эволюции? — Очевидно, да. Но каков механизм этого явления? Это пока остаётся загадкой. Но так ли она неразрешима?
Конечно, если придерживаться старых представлений о происхождении человека и его эволюции, то вряд ли когда-нибудь наука сдвинется с места в этом вопросе. У нас ведь существует привычная избитая схема: бегом от обезьяны через саванну к австралопитеку, питекантропу, неандертальцу, и, наконец, к Homo sapiens. Вроде того как: «Я на дереве сидел, ел банан и вдаль глядел. Вниз летела кожура, вдруг я понял — мне пора. Тут я с дерева сошёл, эволюцию прошёл…». Даже консервативные антропологи уже начали сомневаться в таком примитивном толковании. А тут ещё шведский зоолог Ян Линдблад, автор книги «Человек: ты, я и первозданный», навёл шума в учёном мире, откорректировав теорию Дарвина, где вполне обоснованно доказал, что в своём развитии ещё на раннем этапе, когда появилось прямохождение, человек прошёл водный этап, что и стало в дальнейшем той вехой, которая навсегда отделила человека от его обезьяньих предков.
Но и это не вся правда. Совершенно очевидно, что с дальнейшей эволюцией человеческое общество дробилось на локальные группы с присущими только им особенностями. И эти группы, с прогрессивными, или же атавистическими, признаками сосуществовали на протяжении миллионов лет. Существует совершенно дикая версия о том, что-де каждая новая формация уничтожала предшествующую. Но если следовать такой логике, как объяснить сохранение на Земле казалось бы таких отсталых, но опасных для человека его родичей, как гориллы? А троглодит (т. н. «снежный человек»)? Ведь судя по наличию у него волосяного покрова, его от человека отделяет то же, что и прочих гоминид. А прямохождение (при наличии весьма длинных передних конечностей) лишь свидетельствует о том, что он вступи в параллельную с человеком эволюции сравнительно недавно. Однако, судя по всему, возросшая активность человека перечеркнёт этот шаг нашего меньшого (не по габаритам) собрата. Т. е., я хочу сказать, что в отличие от поспешных заявлений нек. исследователей я придерживаюсь мнения о том, что троглодит — не потомок «деградировавших» палеоантропов-неандертальцев, которым, как уже признано учёным миром, была присуща высокая социальная организация (они умели, в частности, изготовлять довольно сложные орудия, жилища, отправлять культ умерших предков и т. п.), и которые вовсе не были обволошены, подобно троглодиту — а гораздо более древний родственник нашего пращура. Кстати, тот же Ян Линдблад приводит пример того, что человеку, как многим млекопитающим, вовсе не было необходимости вновь обзаводиться волосяным покровом для защиты от холода — его защищает подкожный жир. Он был свидетелем, как аборигенка Огненной Земли (Ю. Америка) с ребёнком переплывала пролив в ледяной воде. Вспомним в этой связи и о наших «моржах». Я могу уверенно назвать научное наименование ископаемого предка троглодита — это гигантопитек Блэка (Gigantopithecus blacki).
Не секрет, что антропологи нередко натыкаются на «неандертальские» признаки среди совр. населения. И это говорит вовсе не о том, что более прогрессивные виды человека смешивались с менее прогрессивными, в частности, неандертальцами, а о наличии пережиточных, не до конца ещё отброшенных природой особенностей. Проще, это означает, что все пра-человеческие и человеческие виды прошли эволюцию в той или иной степени. И именно это — родовое различие — и явилось причиной возникновения человеческих рас. Но не в том понимании, какое придаёт совр. наука этому вопросу, когда утверждает, что всё человечество делится лишь на 3—4 большие расы, а все остальные, более мелкие таксономические подразделения — лишь производные от них. Я как раз считаю, что именно малые расы — вот тот продукт, который был получен природой в результате этого грандиозного действа. Дальнейшее, более мелкое деление — на расовые типы — продукт уже более совр. эволюции, когда из состава малых рас начали выделяться прото-племена.
Но при совр. социальной организации человечество начинает новый виток своей эволюции, когда различные по своему расовому происхождению индивиды дают смешанные типы, в результате чего генная программа не только становится более разнообразной, но и начинает давать сбои, приводящие к генетическим отклонениям, выражающимся через разного рода тяжёлые недуги. И лишь в нек. случаях начинают проявляться нек. т. н. «сверх-нормальные» и «пара-нормальные» качества.
Однако, не следует думать, что в скором будущем всё человечество, как говорил небезызвестный герой Шолохова из «Поднятой целины» Макар Нагульнов, «смешается и станет одинаково смуглявым». Этого не произошло за прошедшие тысячелетия, чему свидетельством довольно замкнутый образ жизни первобытных племён, этого не произойдёт и в обозримом будущем, поскольку расовое и родо-племенное прошлое неистребимо сидит в нас. Каждая раса и расовый тип сугубо консервативны, а в любом обществе подспудно сидит мысль о своей исключительности. И всё-таки задуманная Природой программа будет осуществляться всё с той же методичностью. И этому не может помешать самая избирательная политика расовой сегрегации.
Жизнь не стоит на месте, также как и эволюция. Человек совр. вида (Homo sapiens) — по развитию общей морфологии — появился на Земле, по самым последним данным, не 300 тыс. л. н., а более 2—3 млн. л. н. Понятное дело, что в его 7-млрд. популяции за этот период успели накопиться генетические изменения, зачастую незаметные, тайна которых кроется в морфологии человеческих рас. И не за горами время, когда на смену придёт нечто иное, на нас не похожее внутренне. Нет, конечно, человек не исчезнет с лица Земли, и вряд ли значительно изменится внешне (не учитывая применения технических гаджетов), но что касается внутренних изменений… Уже достаточно давно в обществе известны люди с пара-нормальными способностями, и их становится всё больше и больше, поскольку их гены «кочуют» в мире, передаваясь по наследству из поколения в поколение, но не обязательно проявляясь. Это ещё не сверх-люди, это их предтечи, поскольку 1-м сверх-человеком станет тот, кто соединит в себе все эти сверх-способности. Как только в мире накопится необходимый генетический потенциал, человечество ожидает появление новой формации — сверх-человека, Homo sapientissimus. Где это произойдёт, сказать трудно. Но совершенно очевидно, что место это не будет случайным. Если ещё сохранится деление на страны и общества, то это будет, вероятнее всего, общество с наивысшим потенциалом. Почему? — Прошу обратить внимание, какое значение придаётся сейчас технологии, как интенсивно идёт методичная перекачка «мозгов» в развитые страны Запада, как, впрочем, и лучшего генофонда. Такой потенциал не может быть потерян всуе, генная память постепенно увеличивается, вмещая в себя всё новые качества и показатели.
Но это ещё не вопрос. Основной вопрос перед человечеством встанет тогда, когда оно окажется в роли диких зверей по сравнению с новоявленным хозяином Земли. Что случится с отсталыми тогда Homo sapiens, вольются ли они органично в новое общество, или же их ожидает участь быть занесёнными в Красную Книгу, а то и выселения в глубины Космоса? Ах, если б знать! Но, в конце концов, это вопрос всё-таки будущего…
Как говорит акад. РАН Ю. А. Фомин, и обычные люди за пару лет способны добиться потрясающих результатов. Это их шанс не остаться на задворках прогресса. Для этого нужно лишь одно: долго и упорно учиться. Главная задача сегодня — не вложить в человека максимум знаний, а научить его оперативно получать ту информацию, которая нужна ему здесь и сейчас для каждой конкретной ситуации, из которой складывается наша жизнь. И чем больше правильных решений будет принято, тем гармоничнее она будет. Избежать ошибки, выбрать не единственное, но правильное, решение — и жизнь принимает кардинально иной характер. Т. е. человек, имеющий больше возможностей получать информацию, всегда более удачлив в делах, более счастлив в личной жизни. Он обеспечивает себе преимущество в принятии решений по сравнению с другими людьми.
Можно научиться читать с закрытыми глазами, видеть предметы через препятствие, научиться «слышать» мысли человека и взаимодействовать с ним тактильно на расстоянии — не дотрагиваясь, можно двигать предметы взглядом, зажигать взглядом огонь, можно отрываться от земли, преодолевая гравитацию… Акад. РАН Н. Н. Денисов доказал многое из этого не словом, а делом. Он поясняет: «Волновой сигнал, идущий от объекта, распространяется во все стороны, а не прямо в глаза. Поэтому можно видеть не только ими, но также используя другие органы чувств („видеть“ ушами, носом, пальцами — т. е. с помощью суха, обоняния, ощущений). Когда мы поймём, что первая информация ничем, по сути, не отличается от второй, то поймём и то, что используя второй, нетрадиционный, канал, мы можем видеть гораздо больше, чем обычный человек видит глазами». Чаще всего пара-нормальные способности проявляются в экстремальных случаях. Но совсем не обязательно их ждать.
Если мы будем оставаться такими же, как 10 тыс. л. н., наше жизненное пространство начнёт сужаться. Его потеснят сложные процессы, постоянно происходящие вокруг нас. Когда говорят, что завтра будет легче — не верьте! Завтра будет сложнее! Единственный выход — начать ускоренно развиваться и не отставать от окружающего мира и людей со сверх-способностями.
Живые существа общаются с окружающим миром в гораздо более широком диапазоне, чем человек. Почему они могут, а человек — нет? — И человек может, если очень захочет! Для этого нужен каждодневный труд.
Глава 2 Кто мы? Есть проблема? — Нет проблемы
Согласуясь со своей предыдущей темой (ст. «Сверх-человек. Сегодня ли это уже?»), мне хотелось бы теперь поднять извечный вопрос, пожалуй, самую древнюю мысль человека разумного: откуда мы взялись?
На эту тему не умолкают научные споры, делаются всё новые открытия, зачастую, кажется, перечёркивающие друг друга. И, всё-таки становится очевидно, что тот единственный путь к истине между двумя полюсными взглядами всё ж таки существует, хотя и не везде ещё видны его изгибы и повороты…
Поэтому, представляя на всеобщее рассмотрение свою точку зрения на эту тему, я должен остановиться на последних научных открытиях. Откровенно говоря, у меня не было цели примирить кого бы то ни было, лишь хотелось истины…
В 1987 г. генетики, а именно, амер. биолог Алан Уилсон из Беркли (Калифорнийский университет) совместно с группой университета Эмори, изучив митохондриальную ДНК (мтДНК) 147 человек, представляющих 5 обширных географ. регионов, объявили о своём открытии «генетической Евы». Речь идёт о нашей общей Праматери, чьи гены — одной-единственной женщины — находятся во всех живущих сегодня человеческих существах. В результате этих исследований выяснено, что человек — молодой биологический вид. И что самое интересное — на этом уровне не нашлось никаких межрасовых различий, хотя внешние легко-уловимые различия между расами, казалось бы, свидетельствуют обратное. Парадокс?
Основываясь на этих исследованиях, биологи сделали вывод о том, что совр. тип человека возник где-то между 300 и 200 тыс. л. н., т. е. примерно тогда же, когда происходило становление неандертальцев (палеоантропов). На этом же основании последовал вывод о том, что неандерталец вовсе не является предковой формой совр. человека, поскольку-де митохондриальные гены его «исчезли бесследно» после прихода потомков Евы.
За свои исследования Алан Уилсон заслужил макартуровскую стипендию гения (1986 г.).
Опять же, последние находки палеоантропологов свидетельствуют о том, что планка древности т. н. кроманьонского человека поднимается всё выше и выше. Так, археол. раскопки неподалёку от Сиднея позволили австралийским учёным сделать вывод о том, что человек совр. физического типа появился на Земле (в частности, в Австралии) на 7 тыс. лет раньше, чем считалось ранее: находки из карьера возле р. Непиан датированы 47 тыс. лет., что на 12 тыс. лет древнее для подобных экземпляров в Европе. Группа же франц. и израильских учёных на страницах англ. журнала «Nature» рассказала о том, что человеческие существа, подобные нынешним, были известны на Б. Востоке уже 92 тыс. л. н. Речь идёт о находках в пещере Кафцех/Кафзех, недалеко от библ. г. Назарет. Возрастом 98—96 тыс. лет оцениваются наскальные рисунки во франц. пещере Ля-Марш, изображающие, в частности, лица европ. типов.
В этой связи всё более убидительной становится теория пре-сапиенса, появившаяся в 1970-х гг. и энергично разрабатываемая многими видными з.-европ. антропологами, согласно которой одновременно с неандертальцами, и даже раньше их, существовали-де люди, мало отличающиеся от современных. В частности, речь идёт о находках в Сванскомбе (Англия), Фонтешеваде (Франция), из долины Омо (Эфиопия). Так, известный антрополог Ричард Лики объявил о находке ископаемого черепа неандертальского периода, принадлежащего «существу, которое умело говорить, использовать огонь, мастерило оружие, охотилось, имело свою культуру. Если бы этот парень зашёл сейчас к нам в комнату, вы бы с трудом отличили бы его от наших современников…».
Тем не менее, эта теория не даёт ответа на вопрос, откуда же взялись эти прогрессивные черты у вида, якобы более древнего, чем неандертальцы, вопреки природным законам постепенного эволюционного развития, и почему выжил-де только он, а не не менее развитые, и даже физически более крепкие, неандертальцы. Т. о. эта теория заводит в тупик.
С другой стороны, опять же в журнале «Nature», франц. учёные сообщают, что они нашли останки неандертальцев и их сложные орудия труда, чей возраст составляет приблизительно 36 тыс. лет. Это двойное открытие: оказывается, неандертальцы жили позже, чем считали раньше, а, кроме того, были достаточно разумны. Группы, останки которых были найдены во Франции, имели довольно совершенные инструменты. Например, один из них, предназначенный для резьбы по мамонтовой кости, как до сих пор считалось, использовался только кроманьонцами.
Кроме того, по сообщению «Сан-Франциско кроникл», тель-авивские (Израиль) анатомы обнаружили новые сенсационные свидетельства того, что неандертальцы вполне могли общаться с помощью разговорной речи. Ими была изучена небольшая подъязычная кость возрастом 60 тыс. лет, строение которой доказывает анатомическое сходство неандертальцев и совр. человека: «Практически невозможно найти каких-либо различий».
И, наконец, последнее сенсационное заявление франц. учёного Жерара Люкота, продолжившего исследования в направлении, начатом исследователями «генетической Евы», по сообщению «Сьянс э-ви» (Париж), сделанное в 1991 г., утверждает, что существовал также всеобщий генетический праотец — «Адам».
По последним данным новой научной дисциплины — молекулярной антропологии, ниспровергшей многие понятия классической антропологии, «Адам», прекрасно вписавшийся в схему, разработанную в 1980-е гг., был пигмеем (от греч. «пугмайос» — т. е. «высотой с локоть»). Его наиболее близкими потомками являются, вероятно, совр. пигмеи племени ака, средний рост которых менее 1,5 м.
Т. о. окончательно доказано происхождение всех ныне существующих людей от одного общего корня, и весь вопрос состоит лишь в том, куда же «исчезли» предки расовых групп. В то же время тот же Люкот сравнил гены хромосомы Y с такой же хромосомой шимпанзе — примата, наиболее близкого к человеку, и нашёл, что они находятся практически в таком же сочетании, сходясь в 99%. Давно уже ы сдеан вывод о том, что они произошли от общего предка 5,5—5 млн. л. н.
Теперь пришёл черёд задать вопрос о том, почему, удревняя таким образом человеческую родословную до 6—5 млн. лет, учёные никак не могут отрешиться от классической схемы происхождения совр. человека 200 тыс. л. н.? Возможно, немалую роль здесь играет страх быть обвинённым в расизме? Но наука обязана быть объективной…
Все выводы, сделанные ранее, свидетельствуют о том, что возраст генетических «Евы» и «Адама» вовсе не 200—300 тыс. лет, а не менее 5 млн., о чём и говорит отсутствие различий между нын. расами на митохондрильном уровне ДНК, и что в схему предков нын. человеческих рас прекрасно укладываются различные формы «прото-людей», начиная с австралопитеков, и кончая кроманьонцами. Т. е. я хочу сказать, что все ранее существовавшие пра-человеческие типы, будь то питекантроп рудольфский или яванский, синантроп, неандерталец переднеазиатский или европейский, кафцехский человек, кроманьонец — все в той или иной степени прошли эволюционный путь, явившись той базой, на которой сформировались ныне существующие расы и антропологические типы. Именно этот процесс, происходивший в течение миллионов лет, и стал причиной расовых различий, которые в первую очередь определяются вовсе не цветом кожи (которого мы не можем знать у наших ископаемых предков), не разрезом глаз или особенностями строения скелета и т. п., а, главным образом, формой черепной коробки, различие которой в деталях у представителей одной расы от другой бесспорно, что неизвестно как может отражаться на умственном развитии и аналитических способностях.
Человечество — это дерево, от ствола которого идёт множество ветвей, на которых появляются новые отростки. И всё оно находится в постоянном процессе эволюции. Это можно обрисовать таким образом: когда появляется новая особенность у какого-либо индивида, природа или выбраковывает её в последующих поколениях, или закрепляет в генной памяти, постепенно накапливая в пределах локальной группы. Так начинает проявляться многообразие мира.
Очевидно, постепенно между ныне сложившимися наиболее консервативными замкнутыми расовыми группами накопится такое количество различий, что и впрямь возникнет неск. новых видов, неспособных к скрещиванию. Но это — в теории. На практике же любая, даже самая замкнутая группа испытывает приток генов других групп, что и не даёт произойти окончательному разрыву человечества.
Окончательным подтверждением этому выводу служит истор. факт, повествующий историю дикой женщины-троглодита, прожившей много лет в одном из аулов Абхазии.
Она попала в плен в сер. 19 столетия. По-видимому, пытаясь освободиться, вела себя агрессивно, т. к. её избивали палками и выжги на щеке тавро. Затем её выкупил из неволи сердобольный князь Ачба и посели в своём ауле Тхина, ныне Очамчирского р-на. Ей дали имя Зана. Она жила много лет в этом ауле, в доме селиться не желала, вырыла себе что-то вроде пещеры. С трудом её приучили носить набедренную повязку. Она свыклась со своим положением, выполняла простейшие работы, переносила тяжести, ходила по воду. От молодых абхазов у неё стаи рождаться дети. Вначале они умирали, т. к. она тотчас же после родов носила их омывать, к тому же зимой, в холодный горный ручей. Но потом сердобольные женщины помогли ей воспитать двух сыновей и двух дочерей. Они жили на полных правах вместе со всеми в ауле. От них родились дети — уже внуки Заны. Умерла она в 1880—90 гг. и похоронена на сельском кладбище. Дети её отличались высоким ростом, большей, чем обычно, волосатостью, во всём остальном были такими, как все. В 1930-х гг. этот аул посетил знаменитый скульптор-антрополог м. М. Герасимов, пытаясь найти могилу Заны. Старики-абхазы, помнившие Зану, будучи ещё детьми, несмотря на все старания, не могли найти места, где она была похоронена. Но могилу двух её сыновей обнаружить удалось, и черепа их попали к Герасимову.
Подробное описание внешности Заны даёт Б. Поршнев в своей ст. «В защиту троглодитов». В сокращении это звучит: «… была очень рослая, массивная, широкая. Непомерно большие груди. Высокий толстый зад. Мускулистые руки и ноги, но голень от колен до лодыжек… без всякого утолщения посредине. Пальцы на руках толще и длиннее человеческих. На ногах обладали способностью широко раздвигаться (в т. ч., когда она была раздражена), особенно отодвигался большой палец… Кожа была чёрной или тёмно-серой, всё тело её с головы до ног, и особенно в нижней части, было покрыто чёрно-рыжими волосами, они были местами длиной в ширину ладони, но не очень густы. На лице они были совсем редкие, небольшие, зато на голове, как папаха, возвышалась беспорядочная, свалявшаяся копна совершенно чёрных, жёстких блестящих волос, гривой спускающихся на плечи и на спину… Удивительное было лицо Заны. Оно пугало. Широкое, скуластое, с крупными чертами. Плоский нос со вздёрнутыми большими ноздрями. Выдвинутая вперёд нижняя часть лица — наподобие рыла. Широкий разрез рта, крупные зубы, как-то неестественно выступающий затылок. На широком лбу волосы начинались от самых бровей — лохматых и густых. Глаза имели красноватый оттенок. Но самое ужасное: выражение этого лица было не человеческое, а звериное. Иногда, хотя и редко, Зана рывком, неожиданно начинала смеяться, обнажая свои белые зубы. Никто не заметил, чтобы она улыбалась или плакала…».
К большому сожалению, поведение Заны, историю её жизни в мелочах никто как следует вспомнить не смог. Рождение плодовитых потомков от Заны показывает генетическую близость дикого человека (троглодита) к человеку разумному и представляет исключительный интерес для биологов-генетиков.
Судя по всему, случаи рождения людей с волосами на теле и лице не являются случаями атавизма, как они были объявлены медиками и антропологами, а служат проявлением генной памяти в энном поколении. И это, очевидно, не единственный факт выражения родства с дикими людьми.
А что касается генетических «Адама» и «Евы», так они, как очевидно, были австралопитеками, имевшими именно такой рост. В отношении же объёма головного мозга можно отметить, что он и сейчас значительно варьирует.
Это ещё не конец, до окончательного закрытия вопроса ох как далеко! Учёным предстоит провести ещё огромную работу, для того, чтобы расставить все точки над «i». И, может быть, в скором времени мы узнаем ещё, что-нибудь, что подвинет нас ближе к разгадке собственного прошлого. Ведь говорится же — «не зная прошлого — невозможно уверенно смотреть в будущее». А что касается догм, так не все ли мы уверенно доказали, что «ничто не вечно под луной»? Привычные схемы рухнули, раскрыв перед нами эту простую истину…
Глава 3 Древние гоминиды. Родство с Homo
В свете последних открытий в области палеоантропологии следует произвести ревизию и коррекцию взглядов на происхождение Homo и место его предков в эволюционном ряду среди прочих гоминид, давших начало совр. видам человекообразных обезьян.

Дриопитек

Проконсул
Наиболее древним предком человекообразных (гоминид) считается дриопитек (Dryopithecus, «древесная обезьяна») проконсул, останки которого были найдены в слоях древностью 20±5 млн. лет. Почти современен ему турканапитек (относящийся к роду проконсулов) с оз. Туркана (быв. Рудольфа) из Кении, с укороченным лицом. Ближайшим потомком последнего, очевидно, является совр. гиббон, обитающий в ЮВА — человекообразная обезьяна, наиболее приспособленная к лазанию по деревьям (говоря о гиббоне, следует сказать, что он относится к отдельному семейству — Hilobatinae). В предки гиббона включаются также проплиопитек (Propliopithecus haekeli) — примат среднего олигоцена (30 млн. лет) с изменённым зубным аппаратом, напоминающим аппарат гоминид, и формой челюсти, ближе к человечьей (останки его были найдены в Файюмской пустыне Египта, где некогда простиралась обширная дельта древ. Нила), и плиопитек, имевший руки, которые были короче ног (это как раз удаляет его из предков гиббона), что, очевидно, является свидетельством прямохождения. Проплиопитек, в то же время, считается также предком рамапитека (Ramapithecus punjabicus), размером с собаку, считающегося дальним прародителем человека (хотя до сих пор строение тела его неизвестно), жившим 16—8 млн. л. н. Реконструкция показывает, что у рамапитека было уплощённое лицо, как у гоминид, и близкий к ним зубной аппарат. Останки его обнаружены в Африке, Европе, Индии и Китае. Обитал он в лесах, в саванне не обнаружен. В отличие от него человекообразные обезьяны-вегетарианцы сохранили крупные резцы и длинные клыки на продолговатой челюсти, т. е. различие в зубном аппарата означает также различие в питании. Очевидно, рамапитек начал осваивать ресурсы водоёмов там, где реки были окаймлены лесами. В Китае представители этого вида обитали в болотистом (буроугольном) лесу. Питался он плодами: зубы стёрты также, как у большинства обезьян. Характер стёртостей такой же как и у более позднего австралопитека робустуса и шимпанзе. В его пищу ещё не входили злаки, т. к. он ещё не начал осваивать степную среду.
Ещё один вид из Кении с оз. Туркана — т. н. афропитек, ростом с нын. шимпанзе, с длинным носом, более высокорослый, чем турканапитек, явно претендует на роль совр. носача Борнео (Калимантана) — не уступающего размерами шимпанзе — и носатых обезьян Гималаев (открытых сравнительно недавно). Благодаря своему носу с системой запорных клапанов, самцы носачей (у самок нет такого роскошного «нюхала») способны преодолевать водные препятствия.
Все эти ископаемые виды резко различаются между собой. При этом следует подчеркнуть, что гиббон и носач — обитатели ЮВА, в то время как их ископаемые предки — африканцы. Это свидетельствует если не о большом ареале их распространения в древности, то о миграции из Африки в Азию.
Кроме этих видов, известно большое количество других ископаемых понгид и гоминид: ореопитек, кеньяпитек («кенийская обезьяна»), идентичный рамапитеку, австриапитек («австрийская обезьяна»), испанопитек («испанская обезьяна»), лимнопитек («озёрная обезьяна»), мотопитек («жаркая обезьяна») из Кении, возрастом 7 млн. лет, амфипитек (Amfipithecus mogaugensis, «промежуточная обезьяна» из Могаунга) — один из первых приматов, которые жили на заре возникновения высших обезьян, египтопитек («египетская обезьяна») — из Файюмского оазиса в Египте, размером с гиббона, сочетавшие в своём облике и анатомии черты низших и высших обезьян, весом ок. 10 кг (обе половинки нижней челюсти уже срослись, как у высших обезьян, а зубы напоминают зубы антропоидов, живших 35—30 млн. л. н.: 3 коренных зуба с каждой стороны являются примитивным признаком в отличие от высших обезьян, имеющих по 2).

Носач

Гиббон
В число человекообразных (исключая носачей), кроме гиббона и человека, входят орангутан («лесной человек») — также живущий в ЮВА на о-вах Суматра и Борнео, 5 видов шимпанзе (в т. ч. Pan throglodites и Pan panicus, или карликовый шимпанзе-бонобо), 2 вида горилл (равнинная/саванная и горная, более крупная), а также троглодит («пещерный человек»).

Орангутан
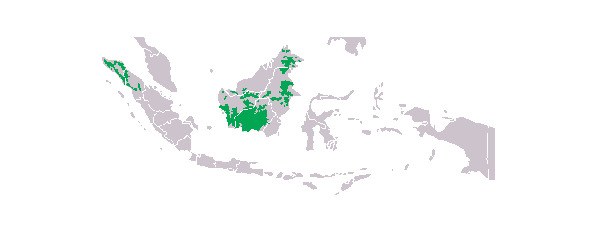
Ареал орангутанов

Бритый шимпанзе
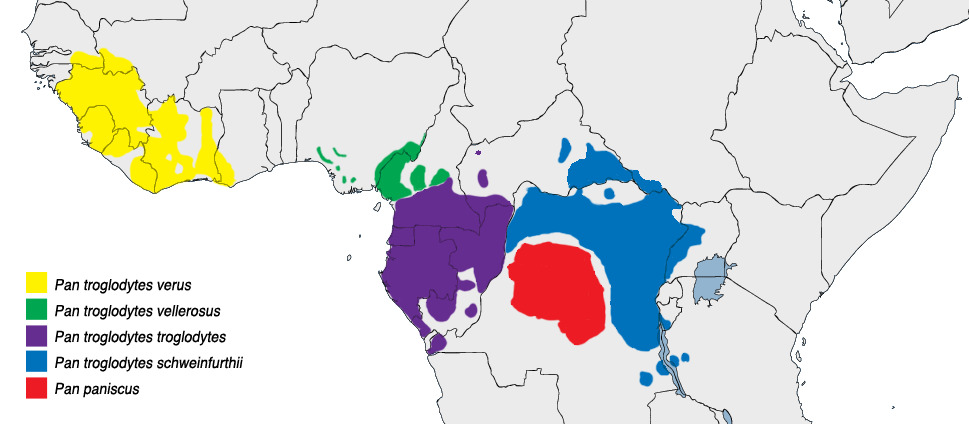
Ареал шимпанзе

Горилла
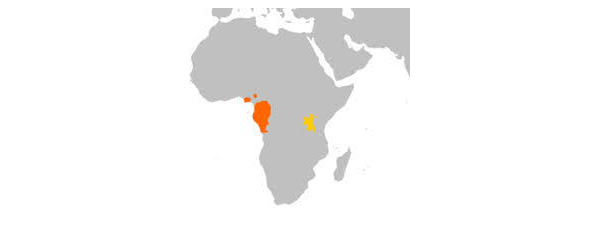
Ареал горилл
По результатам последних исследований выделяют 3 вида орангутанов с 3-мя подвидами в одном из них:
Pongo pygmaeus — Калимантанский орангутан
1. Подвид Pongo pygmaeus pygmaeus (обитает на с.-з. Калимантана)
2. Подвид Pongo pygmaeus morio (с.-в. о-ва)
3. Подвид Pongo pygmaeus wurmbii (ю.-з. О-ва)
Pongo abelii — Суматранский орангутан
Pongo tapanuliensis — Тапанульский орангутан (открыт в 1997 г., классифицирован как отдельный вид в 2017 г.)
Ранее выделялся один вид: обыкновенный орангутан (Pongo pygmaeus) с подвидами калимантанский орангутан (pygmaeus pygmaeus) и суматранский орангутан (Pongo pygmaeus abelii).
Признано неск. подвидов обыкновенного шимпанзе:
Черномордый шимпанзе (Pan troglodytes troglodytes) — в Камеруне, ЦАР, Экваториальной Гвинее, Габоне, Республике Конго и Демократической Республике Конго;
Западный шимпанзе (Pan troglodytes verus) — в Гвинее, Мали, Сьерра-Леоне, Либерии, Кот-д’Ивуаре, Гане и Нигерии;
Pan troglodytes vellerosus — в Нигерии и Камеруне;
Швейнфуртовский шимпанзе (Pan troglodytes schweinfurthii) — в ЦАР, Южном Судане, Демократической Республике Конго, Уганде, Руанде, Бурунди, Танзании и Замбии.
Бонобо (карликовый шимпанзе) — известен давно, но описан как отдельный вид сравнительно недавно — в 1928 г. Несмотря на своё название, по размеру он не меньше шимпанзе обыкновенного, но уступает ему в плотности телосложения. Бонобо встречаются только в Ц. Африке, к югу от р. Конго и к северу от р. Касаи (приток Конго), во влажных лесах Демократической Республики Конго в Ц. Африке.
В нын. время род горилл (лат. Gorilla) относят к семейству гоминид, которое включает и человека. Согласно последним исследованиям (Primate Taxonomy, Colin Groves, 2001. — ISBN 1-56098-872-X) род горилл включает 2 вида с 2-мя подвидами каждый:
Западная горилла (Gorilla gorilla)
1. Западная равнинная горилла (Gorilla gorilla gorilla)
2. Западная речная горилла (Gorilla gorilla diehli)
Восточная горилла (Gorilla beringei)
1. Горная горилла (Gorilla beringei beringei)
2. Восточная равнинная горилла (Gorilla beringei graueri)
Согласно более ранней классификации горилл относили к семейству понгид и выделяли 1 вид с 3-мя подвидами:
Горилла обыкновенная (Gorilla gorilla)
1. Западная береговая или равнинная горилла (Gorilla gorilla gorilla)
2. Восточная горная горилла (Gorilla gorilla beringei)
3. Восточная равнинная горилла (Gorilla gorilla manyema)
Гиббоны (лат. Hylobates) — род человекообразных приматов, обитающих в ЮВА. Один из 4 родов семейства гиббоновые. Раньше был единственным родом в этом семействе, но недавно в категорию родов были также выделены номаскусы, сиаманги и хулоки. Гиббоны являются наиболее разнообразным и широко распространенным родом гиббоновых, обитая на территориях от Ю. Китая (пров. Юньнань) до З. и Ц. Явы. Представители этого рода имеют 44 хромосомы и часто имеют кольцо белого меха вокруг лица.
Род Гиббоны:
Белорукий гиббон (Hylobates lar)
1. Hylobates lar lar
2. Hylobates lar carpenteri
3. Hylobates lar entelloides
4. Hylobates lar vestitus
5. Hylobates lar yunnanensis
Чернорукий гиббон (Hylobates agilis)
1. Hylobates agilis agilis
2. Hylobates agilis unko
Гиббон Мюллера (Hylobates muelleri)
1. Hylobates muelleri muelleri
2. Hylobates muelleri abbotti
3. Hylobates muelleri funereus
Серебристый гиббон (Hylobates moloch)
1. Hylobates moloch moloch
2. Hylobates moloch pongoalsoni
Кампучийский гиббон (Hylobates pileatus)
Карликовый гиббон (Hylobates klossii)
Белобородый гиббон (Hylobates albibarbis)
Гибриды гиббона Мюллера (Hylobates muelleri) и чернорукого гиббона (Hylobates agilis) встречались на территориях Борнео.
По мнению микробиологов (а по мнению генетиков? — авт.), основанном на исследованиях крови, наиболее близко к человеку стоит, якобы, шимпанзе, затем горилла — т. е. африканские обезьяны, и уже затем-де азиатский орангутан; троглодит не рассматривается в виду отсутствия подопытного экземпляра. Однако эволюционно наиболее близким сухопутным видом к человеку является именно орангутан. Более того, последние исследования генетиков в этой области выявили целые участки ДНК, одинаковые как у человека, так и у орангутана. Поэтому утверждение о близости к человеку именно афр. обезьян вызывает вполне понятные естественные сомнения.
Гиббон напоминает человека сглаженной формой черепа, что свидетельствует об их родстве по этому морфологическому показателю, а также плоской грудной клеткой и нек. др. признаками. Шимпанзе больше подходит по строению черепа, размерам конечностей (что не является наследственным фактором). Горилла ближе к человеку по общим пропорциям тела, строению кисти, стопы, таза и нек. др. особенностям. В отличие от афр. обезьян, у орангутана, как и у человека, имеются невысокие бугорки на коренных зубах, сходно устроено твёрдое нёбо, также имеется 12 пар рёбер и вертикальный край лопатки расположен параллельно позвоночному столбу, именно у них растут самые длинные волосы, у них же самые разделённые и разнесённые в стороны молочные железы, одинаковый срок беременности длительностью 270 дней. Если человек имеет какой-то общий с обезьянами анатомический или физиологический, или промежуточный, признак, то чаще всего этот признак оказывается именно с орангутаном, а не с гориллой или шимпанзе. Какова же была исходная форма пра-предка человека?
По расчётам микробиологов, изучавших ДНК, разделение шимпанзе на 2 вида произошло 2,85±0,55 млн. л. н. Разделение шимпанзе и предка человека — якобы 6,6±1,1 млн. л. н., а предков шимпанзе и человека с линией горилл — 9,35±1,65 млн. л. н. Почему не сделано то же для линии человека и орангутана? В этом видны догматы и научная предвзятость, к настоящей науке отношения не имеющие. При этом зафиксирована ископаемая форма шивапитека/сивапитека, родственного орангутану, возрастом 10—8 млн. лет. Строение черепа шивапитека более совершенно в эволюционном плане и более «продвинуто» в сторону человекообразных существ. Останки его найдены в С. Индии, Пакистане и Гималаях. Поэтому возникает закономерный вопрос: кто раз и навсегда доказал происхождение человека в Африке от линии, близкой именно афр. обезьянам, а не в Азии? Если орангутан и родственный ему шивапитек обитали в Азии, это не означает, что они, или преемствующий последнему вид не мог мигрировать в Африку или развиваться в Азии в период, когда начинается кардинальный этап выпрямления позвоночного столба, становления прямохождения (развитый в воде — океане) и хватательной локомоции кисти. Молекулярная биология, на исследованиях которой основывается предварительный вывод о более близком родстве человека и шимпанзе — начинающая наука, и результаты её работы необходимо критически переоценить.
В 2000 г. в Кении были найдены ископаемые останки человекоподобного существа, возрастом 6,5 млн. лет (речь идёт о «Lake Mungo 3» — человекообразном существе, возрастом 6 млн. лет, ходившем на 2 ногах и жившем на берегу озера; однако, структура его ДНК лежит-де далеко за пределами вариаций, свойственных совр. человеку, и, как считается, следовательно, не имела-де продолжения, или, точнее, относилась к другому виду — предкам человекообразных обезьян, возможно, мотопитеков, или же «водных людей» — русалок). В июле 2002 г. в Чаде были найдены останки существа возрастом 7 млн. лет, названного сахелантропом чадским (Sahelanthropus chadensis). Этот вид имел сильно развитые надбровные дуги, но у него практически нет гребня. «Тумай» (в просторечии) вполне может претендовать на роль предка человека. Но он же может быть и предком шимпанзе и родичем орангутана (потомком шивапитека и гиббона).
Интересно, что «водоплавающий» носач обитает в ЮВА. Хотя его предки восходят к В. Африке. Следовательно, здесь могли возникнуть специфические условия, способствовавшие переходу части обезьян к полу-водному, а одного из видов впоследствии — и к водному образу жизни, что имело важнейшие последствия, блестяще описанные шведским биологом Яном Линдбладом в книге «Человек: ты, я и первозданный». Достаточно лишь привести нек. характерные отличия человека от обезьян:
Вертикальная осанка
Короткие руки, длинные ноги
Ступня с непротивопоставляющимся большим пальцем
Рука с противопоставляющимся большим пальцем
Редуцированный волосяной покров на теле
Длинные волосы на голове, волосы под мышками и вокруг половых органов у взрослых особей, иногда борода и усы.
Потовые железы по всему телу
Жировая ткань нового типа, особенно у детей
Большой пенис у мужчин
Большие упругие груди у женщин
Крупный череп, больший объём мозга
Меньшие зубы, особенно клыки
Более плоское лицо
Более высокий нёбный свод, более подвижный язык
Мясистые губы
Выступающий вперёд нос, выполняющий функцию запорного клапана
А также другие отличия.
Из сказанного становится понятно, что к периоду 807 млн. л. н. должно было произойти разделение предка человека с ветвью орангутана. От общего их с гиббоном предка им достались вышеназванные известные про-сапиентные качества, которые затем были востребованы в ходе эволюции. Т. о. сходство человека с афр. обезьянами в характеристиках крови ещё ни о чём не говорит, а если и говорит, то. Очевидно, только то, что шимпанзе также родственны общему предку человека и орангутана — шивапитеку, преемствуя последнему как и человек. При этом почему-то традиционно считается, что ископаемых форм афр. обезьян в Азии нет. Но при всём наличии ископаемого материала таковые не признаются и для Африки западнее Большого Рифта, кроме р-на оз. Чад.
В Африке известны ископаемые формы, делящиеся на 2 группы: 1. плезиантропа (Plezianthropus transwaalensis) или австралопитека африканского/протметеева (Australopithecus africanus/prometheus), возрастом 2,7—2,2 млн. лет 2. парантропа грацильного (Paranthropus grassideus) и парантропа массивного (Paranthropus robustus), известного также как зинджантроп/австралопитек массивный/бойсей (Zinjanthropus/Australopithecus robustus/boisei), возрастом 2,1—1 млн. лет, отличающихся крепким/дюжим телосложением и мощным зубным аппаратом.
Совр. палеоантропология условно разделяет австралопитеков на 3 группы:
ранние австралопитеки (7,0—3,9 млн. л. н.)
грацильные австралопитеки (3,9—1,8 млн. л. н.)
массивные австралопитеки (2,6—0,9 млн. л. н.)
Ранние австралопитеки включают в себя роды:
Australopithecus — Australopithecus anamensis (Leakey, Feibel, McDougal et Walker, 1995)
Sahelanthropus tchadensis (Brunet at al., 2002)
Orrorin tugenensis (Senut, Pickford, Gommery, Mein, Cheboi et Coppens, 2001)
Ardipithecus ramidus (White, Suwa et Asfaw, 1995).
Грацильные австралопитеки включают в себя следующие виды:
Australopithecus afarensis (Johanson, White et Coppens, 1978)
Australopithecus bahrelghazali (Brunet, Beauvilain, Coppens, Heintz, Moutaye et Pilbeam, 1996)
Australopithecus africanus (Dart, 1925)
Australopithecus garhi (Asfaw, White, Lovejoy, Latimer, Simpson et Suwa, 1999)
Australopithecus sediba (Berger, 2010)
Kenyanthropus platyops (Leakey, Spoor, Brown, Gathogo, Kiarie, Leakey et McDougalls, 2001)
Последняя группа из-за своей специфической анатомии выделяется в отдельный род — Paranthropus, насчитывающий 3 вида:
Paranthropus aethiopicus (Arambourg et Coppens, 1968)
Paranthropus boisei (Leakey, 1959)
Paranthropus robustus (Broom, 1939).
Существует ещё неск. спорных видов, которые могут быть отнесены к группе австралопитеков, однако это выходит за рамки настоящей статьи.
Анамский австралопитек (лат. Australopithecus anamensis) — ископаемый вид рода австралопитеков, обитавший в В. Африке ок. 4 мн. л. н. Это наиболее древ. из видов австралопитеков, известных науке.
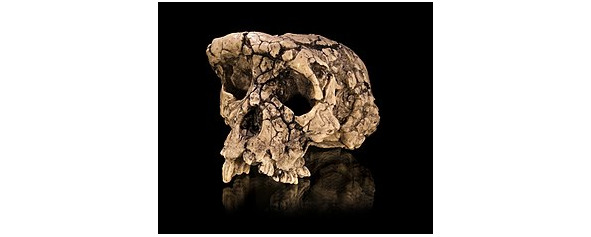
Сахелантроп (лат. Sahelanthropus) — род гоминид, представители которого жили в позднем миоцене (7 млн. л. н.). В роде выделяют один вид — сахелантроп чадский (Sahelanthropus tchadensis). Описан по находкам, сделанным на с.-з. республики Чад (пустыня Дьюраб, Торос-Менелла) в 2001 г.: черепу Тумай и фрагментам ещё неск. особей. По классификации первооткрывателей является самым древ. из известных представителем гоминин.
В р-не оз. Чад, кроме сахелантропа, найдены останки одного из австралопитеков, названного чадантропом (Chadanthropus), который, видимо, преемствует сахелантропу.
Оррорин (лат. Orrorin) — вымерший род афр. больших человекообразных обезьян. В популярной литературе известен как «Человек Тысячелетия» («Millennium Man»). Единственный вид в роду орроринов — Orrorin tugenensis или Praeanthropus tugenensis. Считается одним из древнейших известных предков гоминин и, возможно, совр. людей. Название рода Orrorin переводится с местного языка как «первый человек». Видовое имя tugenensis происходит от названия холмов Tugen Hills (Кения) в месте, где оррорин был открыт. Останки залегали между 2-мя слоями вулканического пепла, благодаря которым датируются относительно точно: между 6,1—5,8 млн. л. н., в эпоху миоцена.
Ардипитек (лат. Ardipithecus) — древ. род гоминид, который жил в раннем плиоцене прим. 4,4 млн. л. н. Из-за сходства зубов с зубами австралопитека считается нек. исследователями древней. предком человека. Кроме того, в стопе ардипитека имеется добавочная малая берцовая кость, которая присутствует в стопе человека, но отсутствует в стопе других совр. гоминид. Точно не установлено, мог ли он ходить вертикально. Иногда ардипитеков относят к ранним австралопитекам. До сих пор было описано только 2 вида: Ardipithecus ramidus и Ardipithecus kadabba. Последний вначале считали подвидом A. ramidus, но по форме зубов, недавно обнаруженных в Эфиопии, определили как особый вид. Все останки найдены в В. Африке, где, видимо, когда-то находилась и родина совр. шимпанзе, которые, как и гориллы, заселили З. Африку.
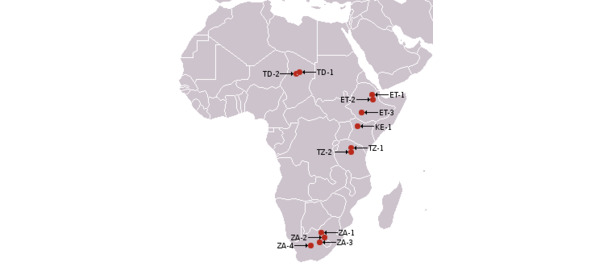
Места находок
Афарский австралопитек (лат. Australopithecus afarensis) — вымерший вид австралопитека раннего, живший ок. 4 млн. л. н. в пустыне Афар (Эфиопия), в местности Хадар. Почти нет сомнений, что до того, как афарские австралопитеки «вымерли» 3,5—2,5 млн. л. н., от них прямо или косвенно произошли другие австралопитецины и род Homo. Афарский австралопитек — самый маленький вид австралопитеков, прямоходящий (хотя ещё не произошла адаптация к поной двуногости: ходил-де, переваливаясь, на неск. согнутых ногах, а с помощью длинных рук (мощная плечевая кость), якобы лазал по деревьям (или же был мощным пловцом), питаясь-де фруктами и корнеплодами), в целом сходен с видом т. н. Homo habilis. Он, вероятно, имел тёмную кожу и был покрыт волосами. Рост, как и у Homo habilis, тоже небольшой — 1—1,5 м, масса тела — ок. 30—60 кг. Этот вид отличала т. н. прецизионная хватка, каковой обезьяны не наделены. В зубном аппарате видны элементы, из которых затем могли развиться крупные зубы, как автралопитеков, так и менее мощные линии Homo. Оуэн Лавджой отмечает, что лёгкий изгиб плюсневых костей говорит о приспособлении для ходьбы по рыхлой, песчаной почве, т. е. он был привязан к реке (р. Аваш, впадавшая в ныне высохшее озеро). Мозг афарского австралопитека не отличается большими объёмами (~380—430 см куб., что немного больше, чем у шимпанзе). Самцы были на 60—70% массивнее самок (такое соотношение нередко у горилл). По мнению Дона Джохансона, представляют тот же вид, что и гоминиды из Лаетоли (Кения), где отмечены следы прямоходящих гоминид возрастом 3,6 млн. лет. Р. Лики отмечал, что зубы и челюстные кости очень похожи на зубы и кости рамапитека: «Пожалуй, можно считать поздним рамапитеком». Здесь же обнаружены останки гоминида, относящегося, по видимому, к этому же виду, ростом ок. 130 см, с выраженными надбровными дугами и низким покатым лбом. Передние зубы-резцы были почти в 2 раза длиннее, что делало его похожим на совр. павианов. Как отмечалось в журнале «Nature», морфология афарского вида очень напоминает совр. карликовых шимпанзе-бонобо, чему, как будто, соответствует и возраст находки. Впрочем, у бонобо более мощные кости таза, чем у афарского австралопитека. И образ жизни их преимущественно древесный. Объём же мозга рядового шимпанзе лежит в пределах 320—480 см. куб. На оз. Баринго (Кения) обнаружен вид австралопитека табаринского, возрастом 5—3,6 млн. лет, по всем признакам близкий афарскому виду.
Бахр-эль-газальский австралопитек (лат. Australopithecus bahrelghazali) — предположительно, самост. вид австралопитека, живший ок. 3,5—3,0 млн. л. н. Это самая западная находка австралопитека в Коро Торо на территории древ. речного русла Бахр-эль-Газаль на юге адм/региона Борку в Чаде.
Сахелантроп
Ардипитек

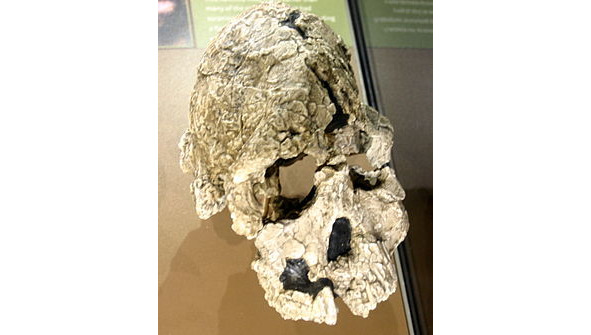
Череп самки африканского австралопитека (плезиантропа)
Африканский австралопитек (лат. Australopithecus africanus) — вымерший вид австралопитека, живший ок. 3,5—2,4 млн. л. н. В отличие от афарского австралопитека имел более обезьяноподобный скелет, но более объёмный череп. Центральное расположение большого затылочного отверстия, через которое спинной мозг соединяется с головным, говорит о прямохождении. Почти нет надбровных дуг. Более грацильный (стройный) вид, в отличие от парантропов. Основные местонахождения останков этого вида — известняковые пещеры Ю. Африки: Таунг (1924), Стеркфонтейн (1935), Макапансгат (1948), Глэдисвэйл (1992). Во многом строение скелета афр. австралопитека имеет множество сходств с его в.-афр. родичем — афарским австралопитеком е. Например, у обоих видов длина рук неск. превышала длину ног. Однако детали строения черепа у афр. австралопитека более близки к человеческим. Плезиантропы были ростом с совр. бушмена (полумонголоида-полунегроида), прямоходящие, с зубной системой, близкой синантропу. Останки их найдены в Ю. Африке. Период их существования приходится на тот же период, что и мегантропов. В эту же группу включается и автралопитек прометеев, найденный в Ю. Африке и Кении, рядом с которым отмечены следы огня (откуда и название). Плезиантропы-автралопитеки могут являться предками различных видов шимпанзе. Для них отмечено, что пясть относительно длиннее, а пальцы короче, чем у совр. человека.


Австралопитек седиба
Кениантроп
Австралопитек гархи (лат. Australopithecus garhi) — вымерший вид австралопитеков, живший ок. 2,5 млн. л. н. Представители этого вида являются возможным «переходным звеном» от рода австралопитеков к роду людей. Относятся к группе грацильных австралопитеков, т. е. обладают хрупким, не массивным телосложением. Удивительность этого вида заключается в том, что вместе с останками были найдены примитивные орудия, напоминающие олдувайскую технологию, и датируемые возрастом 2,5—2,6 млн. лет.
Австралопитек седиба (лат. Australopithecus sediba) — вид австралопитеков, известный по остаткам, возраст которых оценивается в 1,95—1,78 млн. лет (плейстоцен). 2 неполных скелета этого вида были найдены при раскопках в пещере Малапа на территории «Колыбели человечества», в Ю. Африке. Объём мозга — 420 см куб.
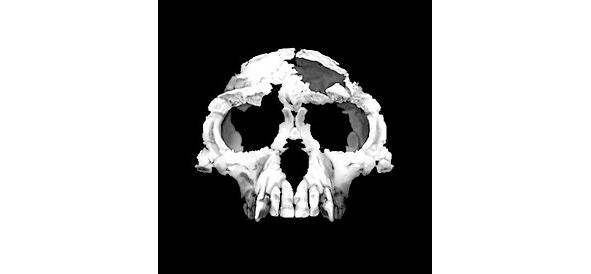
Кениантроп (лат. Kenyanthropus) — древ. род гоминид, живший в плиоцене ок. 3,5—3,2 млн. л. н. Останки кениантропа были обнаружены на берегу оз. Туркана (быв. Рудольфа). По мнению первооткрывателей, не австралопитеки, а именно кениантроп является прямым предком человека. С лат. Kenyanthropus platyops означает «плосколицый кенийский человек». Кроме вида K. platyops, Лики относит к роду Kenyanthropus ещё и вид Homo rudolfensis. Другие же антропологи классифицируют кениантропа как отдельный вид австралопитеков — Australopithecus platyops или интерпретируют его в качестве одного из индивидов австралопитека афарского. В кенийском местонахождении Ломекви 3 на западном берегу Турканы, недалеко от места находки кениантропа, в 2015 г. были найдены и древнейшие в мире каменные орудия возрастом 3,3 млн. лет, что на 700 тыс. лет старше, чем орудия из эфиопского местонахождения Гона.
В С. Индии также найдены фрагменты скелета какого-то вида австралопитека, но относятся ли они к предкам человека или какому-то другому виду гоминид — неясно.
Эфиопский парантроп (лат. Paranthropus aethiopicus) — ископаемый вид рода парантропов («массивных австралопитеков»), обитавший в В. Африке ок. 2,5 млн. л. н. Это наиболее древний из парантропов, известных науке. Эфиопского парантропа не следует путать с другим таксоном, имеющим сходное название — Australopithecus africanus aethiopicus (австралопитеков из местонахождения Хадар (р-н Афар, Эфиопия)). В настоящее время эти хадарские находки обычно классифицируются не как самостоятельный вид или подвид, а как представители афарского австралопитека.
Парантроп Бойса/бойсей (лат. Paranthropus boisei, устар. зинджантроп [syn. Zinjanthropus boisei]) — самый массивный вид парантропов, обнаруженный в 1959 г. в в.-афр. ущельях Олдувай, Кооби-Фора, Локалеи и ряде других. Они датированы 2,5—1 млн. л. н. Вплоть до 1972 г. считался «несомненным австралопитеком». Данный вид обитал в В. Африке в эпоху плейстоцена примерно от 2,4 до 1,4 млн. л. н. Парантроп Бойса — наиболее массивный из всех австралопитеков: характерны огромные моляры, тяжёлые челюсти, которые, однако, не выступают вперёд, большие костные гребни, служившие для прикрепления жевательных мышц. Общая массивность сложения была, видимо, также очень большой — при росте 1,2—1,6 м парантропы Бойса весили от 40 до 90 кг. Взрослые самцы были в среднем крупнее самок, как это было практически у всех австралопитековых видов. Мужчины весили приблизительно 49 кг и их рост был приблизительно 1,37 м, в то время как женщины весили приблизительно 34 кг и были ростом 1,24 м. Парантропы Бойса были полностью прямоходящими. Объём мозга довольно мал, 400—500 см куб.
Парантроп массивный (Paranthropus robustus, лат. буквально — массивный парантроп) — вид ископаемых высших приматов, обнаруженный в Ю. Африке в 1938 г. Останки были датированы периодом от 2 до 1,2 млн. лет. Считаются родственной австралопитекам тупиковой ветвью эволюции человека. Объём головного мозга примерно 520 (530—550) см куб. Вид отличался огромным костяным гребнем на черепе, идущим от затылка ко лбу, для крепления жевательных мышц. Выступающие вперёд скулы также свидетельствуют о развитой жевательной мускулатуре. Паратроп робустус имел руку, более похожую на человечью, где большой палец противопоставляется всем остальным. Походка была отртогнатной. Парантроп бойсей имел такие огромные челюсти и зубы, что ему дали прозвище «щелкунчик». Опираясь на разные данные, палеоантрополог тим Уайт попытался реконструировать его облик: сходство с маленькой самкой гориллы поразительно.
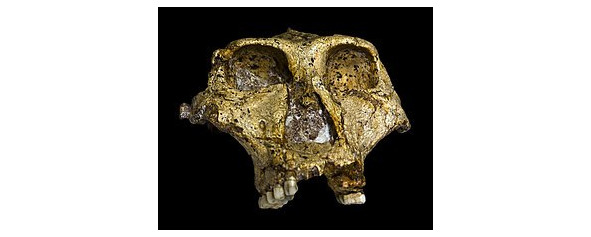



Реконструированный облик парантроп Бойса
В отличие от более грацильных видов плезиантропов-автралопитеков, парантропы отмечаются в Кении и Танзании. Объём их мозга сравним с объёмом мозга горилл — средний составляет 585 куб. см, наибольший известный — 752 куб. см. Т. о. можно рассматривать парантропов и африканских мегантропов как предков горилл.
Известно, что в Азии (С. Индия, Ю. Китай), в ЮВА (о. Ява) и Африке найдены ископаемые останки т. н. рамапитековых обезьян — соответственно, гигантопитека Блэка (Gigantopithecus blacki), мегантропа палео-яванского (Meganthropus paleojavanicus) и мегантропа африканского (Meganthropus africanus), по габаритам не уступавших габаритам гориллы (были даже крупнее), с объёмом мозга 430—550 куб. см, которые, очевидно, вели исключительно наземный образ жизни по преимуществу с вегетарианским питанием, о чём говорит наличие у них гигантских челюстей и зубов. Ранние из них относятся к периоду 2,7—2,2 млн. л. н. Наиболее поздние (Ю. Китай) фиксируются-де 500—300 тыс. л. н.

Реконструкция Gigantopithecus blacki
Кроме австралопитеков нам не известны обезьяны, которые вели исключительно наземный образ жизни (а кто сказал, что и человек вёл его?), и должны-де были неизбежно приобрести относительно ортогнатную походку (каковы должны были быть условия, их на это подвигнувшие, кроме водного образа жизни? — авт.), в связи с чем соответственно изменялась форма стопы в сторону человечьей (скорее, она изменялась в сторону ласта :) — авт.). По своим параметрам с гигантопитеком и мегантропами сходны совр. виды гориллы и троглодита. Троглодит ортограден, наподобие Homo sapiens и массивнее, чем его афр. родичи-гориллы, так и не приобретшие прямохождение, и к тому же тёмнопигментирован (по ряду свидетельств), как и негро-австралоидные народы, хотя зачастую имеет рыжеватую или каштановую шерсть. Естественно, этот вид не может относиться к каким-либо неандертальцам, как это предполагал даже ряд антропологов, или даже питекантропам, не только потому что обволошён и имеет на голове «гребень», как у самцов горилл, но и потому что не владеет речью.
Т. о. версия о том, что к востоку от Великого Африканского Рифта жили ископаемые предки человека и не селились предки афр. обезьян, оказалась неверна, получив серьёзную «трещину».
Человек умелый (лат. Homo habilis) или презинджантроп — вид ископаемых гоминид, высокоразвитый австралопитек. 1-й экземпляр обнаружен в ноябре 1960 г. в ущелье Олдувай в Танзании. Вместе с костями саблезубого тигра смилодона нашли стопу, пяточную кость, ключицу и обломки черепа. Череп, как установили потом, принадлежал ребёнку 11—12 лет. Судя по строению стопы, гоминид был прямоходящим. Находка была описана в 1964 г. как новый род гоминидов — презинджантроп (Prezinjanthropus). Позднее классификация была пересмотрена в пользу видового статуса в роду Homo. В дальнейшем находки представителей данного вида были сделаны в Кооби-Фора, Сварткрансе и других местах В. и Ю. Африки.
Человек умелый, судя по найденным останкам, датирующимся 2,8—2,5 млн. л. н., существовал более полумиллиона лет. Объём мозга человека умелого — 600—700 (а в отдельном случае — до 775) см куб., что намного больше, чем у типичных австралопитеков. Однако, несмотря на больший, чем у прочих австралопитековых, объём мозга, он имел чрезвычайно малый рост, составлявший до 1,2 м и сближавший его с афаро-табаринским видом, длинные руки, доходившие почти до колен и почти равные длине ног, массу тела — ок. 40—50 кг. Его лицо имело архаичную форму с надглазничными валиками, плоским носом и выступающими вперёд челюстями. Голова человека умелого стала более округлой формы, чем у австралопитеков; мозг также стал крупнее, хотя составлял всё ещё лишь половину мозга современного человека. Он также отличался от австралопитеков строением таза. Отличался он от австралопитековых и строением черепа — череп расширен в подглазничной и теменно-затылочной областях. Размер зубов уменьшается. Структура кисти Homo habilis мозаична и сочетает в себе как прогрессивные черты, так и следы адаптации к лазанью по деревьям. У человека умелого первый палец стопы не был отведён в сторону, а, как у всех последующих представителей рода Homo, в том числе Homo sapiens, располагался вместе с другими пальцами. Это означало, что нога его была полностью приспособлена только к двуногому передвижению. Челюсти были менее массивные, чем у австралопитека; кости рук и бёдер кажутся более современными, а ноги имели вполне «современную» форму. У особей человека умелого был заметный половой диморфизм — самки имели более широкие бёдра по сравнению с самцами.
KNM ER 1805, возможно, является гибридом раннего Homo и массивного австралопитека.
Все попытки объявить презинджантропа предком древнейшего человека — Homo erectus — несостоятельны, поскольку последний ростом и телосложением практически не отличается от совр. человека, причём существовал гораздо раньше презинджанторпа (до 2,5 млн. л. н.). Учёные из университета Саймона Фрейзера делают вывод, что человек умелый вышел из Африки до появления человека прямоходящего, став предком человека флоресского.
Как видно из вышесказанного, предок шимпанзе нашёл неск. иную экологическую нишу, как нашли свои ниши вегетарианцы-гориллы и всеядные троглодиты (настоящие троглодиты, в отличие от шимпанзе) с мощными челюстными аппаратами — хозяева не только тропического леса, но, как видим, и тайги.
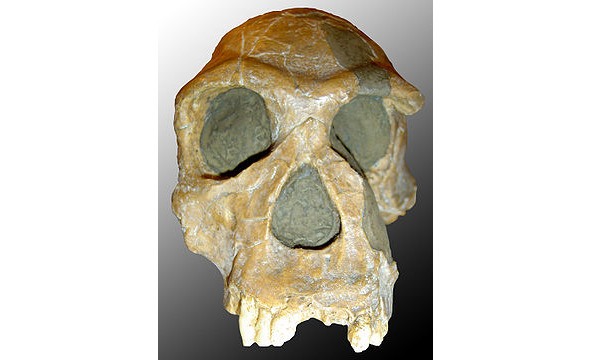
Человек умелый
В 1967 г. международная экспедиция в долине р. Омо в Эфиопии открыла костные остатки, относящиеся к австралопитековым. Вместе с ними были обнаружены и орудия труда — обработанные камни-чопперы. Позже каменные орудия труда возрастом 2,6 млн. лет были найдены в эфиопском местонахождении Гона и возрастом 3,3 млн. лет — у оз. Туркана, недалеко от места находки кениантропа. Т. о., вопрос возникновения орудийной деятельности остаётся открытым.
Нек учёные приписывают виду Homo habilis способность на низком уровне обрабатывать камни (что, как будто, было присуще и парантропу, и что, якобы, является характерной чертой совр. троглодита, руки которого значительно длиннее человеческих и достигают коленей, но сходны по строению кисти). Похоже, однако, что данные способности не были достаточно осознанными действиями, т. е. трудом в прямом смысле этого слова, а являлись, скорее, инстинктивными, вызванными окружающими условиями, т. е. находились на том же подсознательном уровне, что и «трудовая деятельность» пчёл и муравьёв. Точно также морская выдра (калан) использует камни для разбивания скорлупы моллюсков, точно также птицы подражают действиям и речи человека. Т. е. идёт процесс подсознательного копирования некогда увиденных действий (в процессе воспитания).
Исходя из этого посыла, мы должны признать в корне ошибочной марксистскую доктрину (точнее, доктрину Ф. Энгельса) о том, что «труд сделал из обезьяны человека». Считается, что труд начинается с изготовления орудий. Однако, судя по данным археологии, примитивные орудия изготавливали австралопитецины — предки человекообразных обезьян — и троглодиты.
Во-первых, прямохождение появилось у гоминид вследствие причин, связанных с переменой среды обитания и образа жизни, а вовсе не из-за «трудовой деятельности». Во-вторых, то же можно сказать и об увеличении объёма мозга на ранних этапах: поскольку вновь сменив среду обитания, предок человека вынужден был приспосабливаться к новым условиям, превратившись в результате всех предпосылок в Великого Охотника, и, соответственно, увеличил свой белковый рацион. Т. о. не труд создал из обезьяны человека, а охота (хотя, в известном смысле, это тоже — труд). Предок человека сумел занять верхушку биологической пирамиды, только став Великим Охотником. Примеров из истории предостаточно: динозавры достигли доминирующего положения в животном мире, заняв верхушку пищевой пирамиды и как бы остановились в своём развитии (например, дейнонихус — один из самых свирепых и совершенных универсальных хищников всех времён, практически не нуждался в своём совершенствовании — это была идеальная машина для убийства, что, как будто, и является закономерным итогом эволюции), если бы 65 млн. л. н. метеорит, упавший в р-не Мексиканского залива (а его осколки — в других местах), не нарушил этой идиллии, изменив условия развития животного мира на Земле, и создав предпосылки для развития новых видов. Цепочка климатических и природных колебаний и изменений, взаимосвязанных между собой, привела к появлению ветви гоминин, отделившихся от общего гоминидного ствола.
*
Как уже говорилось, архантропы (Homo erectus — человек прямоходящий) — древнейшие люди, появляются свыше 2 млн. л. н. как бы «внезапно» и в Африке (Кения), и в Азии (о. Ява) (последние находки в Якутии (Диринг-Юрях) как будто свидетельствуют о том, что уже 2—1,8 мн. л. н. на севере также жили древнейшие люди; однако, эти находки ограничиваются чопперами). Т. о. промежуток между 7—2 млн. л. н., в который происходила коренная эволюция предка человека, нам практически неизвестен. Единственно бесспорно установленный факт — что человек в своём развитии, в отличие от других гоминид, прошёл водную стадию, что сделало его более мобильным и открыло доступ ко всем уголкам света. Эта стадия была неожиданно прервана, поскольку человек так и не стал единственным полностью водным приматом, но причины этого нам неизвестны, и человек появляется в палеонтологической картине уже как Великий Охотник, использующий жильё, одежду, ловушки и копьё, а также примитивные каменные орудия, а позже — и огонь. Понятно, что сферой его становления были водные бассейны, реки и прибрежные области. Т. о. останки предков человека. Относящиеся к периоду их водной стадии, нам неизвестны, и находка их зависит от правильного направления поисков. Период водной стадии укладывается в отрезок между 5—2 млн. л. н., т. е. на конец плиоцена, когда огромные лесные массивы сжались, уступив место степям (а позднее и пустыням типа Руб-эль-Хали или Сахары, возникшим, однако, по другим причинам), высохли многие водоёмы, поскольку ближайший известный ископаемый предок человека — шивапитек — известен ещё 5—4 млн. л. н. в С. Индии. Т. о. возможно не только рептилии третичной фауны (крокодилы/аллигаторы) и гиппопотамы — свирепые озёрно-речные хищники — вынудили человека покинуть обжитые водоёмы, но и сильнейшая засуха. Те же группы, которые осваивали моря, дали начало нын. русалкам (вид sireni erit). Переход же к сухопутному образу жизни, переселение в другие регионы, и, как следствие, освоение человеком лесов и саванн привели к становлению охоты и возникновению новой формации. Этот последний процесс, как думается, проходил в период между 3 и 2 млн. л. н.
Отделение гоминин в новую ветвь (точнее, вид) от прочих гоминид, точнее, от предковой для гоминин формации шивапитека, близкого азиат. орангутану, могло произойти в р-не Красного моря, отделяющего Африку с большим количеством древ. гоминид от Азии (Аравийского п-ова), где таковых не найдено на противоположном, азиатском, берегу, ныне пустынном (аналогичные пустынные берега и у Персидского залива, но ведь в какой-то период они были зелёными! — авт.). Красное море обладает всеми необходимыми условиями для формирования водного вида гоминид: обширные водные пространства, тропический климат с богатой растительностью и плодами, моллюски, ракообразные, рыба и пр. (наличие моллюсков и ракообразных было исключительно важно для формирования хватательной локомоции кисти и появления каменных орудий, необходимых водному примату для разбивания панцирей после захвата). В тот период Красное море представляло собой, возможно, уже открытый водоём, из которого и началось распространение популяции уже полностью перешедших к водному образу жизни прото-русалок.
Переселенцы же из этого региона (Эритреи), ставшего засушливым, адаптировались к сухопутному образу жизни, положив начало виду Homo erectus.
Почему же формирование гоминин должно было происходить только в одном водном бассейне, а не в нескольких? — Потому что в т. сл. существовала бы не одна, а уже неск. предковых форм человека, возможно, не способных к гебридизации и стоящих на разных уровнях развития, связанных с водным этапом. К тому же климатические условия в разных регионах могли изменяться по-разному, что неодинаково отразилось бы на дальнейшей эволюции популяции гоминин.
В наст. момент больше всего вопросов относится к теме происхождения расовых ветвей человека: как произошло разделение единой предковой популяции Homo erectus на расовые стволы, почему реликтовая койсанская раса-эндемик, являющаяся предковой по отношению к монголоидной, ныне занимает юго-запад Африки, а древние негроидные популяции распространены в Австронезии и на ю.-азиат. о-вах?
Пора прекратить говорить о том, что предок человека «спустился с дерева» и встал на две ноги, без чего ему, якобы, было не выжить в саванне, куда он вышел — как самоубийца — для чего-то из леса, богатого всем необходимым для существования обезьяны. На самом деле передвижение на 4-х конечностях, наоборот, давало ему преимущество в скорости, и он бы им воспользовался, отказавшись от перспективы «освобождения передних конечностей» для, якобы, «созидательного труда», если бы предок его уже не бы прямоходящим, каковым стал достаточно давно, обитая в озёрно-речной и приморской местности. При достаточной массивности, гоминиды, отделившиеся от линии гиббонов, перешли к полу-наземному образу жизни. Это т. н. стадия австралопитековых — почти прямоходящих, и даже прямоходящих приматов, к каковым можно причислить шимпанзе, гориллу, орангутана и троглодита.
Следует считать ошибочной и теорию о том, что с появлением новых, более прогрессивных, формаций, исчезают предыдущие в результате их, якобы, уничтожения потомками (любопытно было бы посмотреть на этот процесс и автора этого пассажа — авт.). Вряд и это отвечает сколько-нибудь реальной жизни: скорее, следует говорить о процессе внутривидовой гебридизации. До появления архантропов (древнейших людей) в мире не существовало хищных гоминид, которые бы могли уничтожать себе подобных, а с появлением Великого Охотника его охотничья деятельность была переключена главным образом на третичную мегафауну. И совсем уж смехотворной кажется «классическая» версия о том, что презинджантроп (Homo habilis) ростом «метр с кепкой» мог являться предком Великого Охотника — человека, и ок. 1 млн. л. н. «уничтожить» дюжего и массивного зинджантропа (парантропа) и мегантропа африканского — предков горилл и троглодита.
Глава 4 Происхождение рас. Центры расообразования
В науке в целом существует 2 разных взгляда на происхождение человека — моно-антропоцентризм, т. е. происхождение из одного центра, и поли-антропоцентризм, т. е. происхождение его из неск. центров. Однако в каждой из этих (довольно многочисленных) теорий есть взаимоисключающие моменты, что ставит под сомнение общую идею и играет на руку креацианистам. Нечто среднее между этими теориями представляет т. н. дицентризм. Чем же характерны эти теории? — Моноцентристы в целом настаивают на начале разделения человечества на расы в кроманьонскую эпоху (не учитывая того факта, что уже ок. 100 тыс. л. н. в Палестине фиксировались палеоантропы группы Схул с расовыми признаками и черепной морфологией, приближенной к неоантропной). Полицентристы же возводят происхождение их ещё ко временам архантропров (Homo erectus), опираясь на данные морфологических исследований.
Крайняя полицентрическая точка зрения, считающая 3 вида человекообразных обезьян предками 3-х человеческих рас дика, смешна и нежизнеспособна, поскольку предполагает одинаковую тенденцию в определённом эволюционном направлении и одинаковую динамику изменения морфологии этих гоминид без учёта специфических особенностей окружающей их природы, в то время как известно, что всё человеческое сообщество одинаково отделено от гоминидного мира рядом генетических показателей, сходных для всех ныне существующих людей. Т. е. на определённом этапе развития гоминидов от них отделилась ветвь, послужившая в дальнейшем основой формирования человечества.
Нельзя всерьёз воспринимать и «классическую» моноцентрическую теорию, утверждающую, что, якобы, каждая новая формация «уничтожала» предыдущую, или же они в той или иной степени распространённости продолжали существовать до появления именно человека совр. вида — Homo sapiens sapiens, поскольку подобные взгляды не учитывают существующие до сих пор виды человекообразных обезьян и отрицает т. о. теорию эволюции.
В конце концов, нельзя утверждать наверняка того, что мы не в состоянии проверить и подтвердить, тем более теории, основанные на чрезвычайно зыбкой почве. Иногда маленький штрих способен испортить всё произведение. Так, например, т. н. Homo habilis (пре-зинджантроп), предок карликовых шимпанзе-бонобо, имевший довольно примитивное строение по сравнению с нек. более ранними и совр. ему австралопитеками, но обладавший большим объёмом мозга и живший ещё в начале эпохи архантропов, на самом деле подтверждает вывод о том, что и наиболее примитивные формы продолжали сосуществовать, эволюционируя, с другими более поздними и более «прогрессивными» видами. Совершенно естественно, что наиболее древние индивидуумы этого вида имели мозг, не превышавший по своему объёму средний показатель обезьян.
Каковы трудности палеоантропологии в формировании теории антропогенеза скажет любой антрополог: фрагментарные одиночные ископаемые находки не позволяют судить о многообразии древних антропологических форм и их популяций, и даже о т. н. «средних» индивидуумах той или иной группы, но в целом складывается общая картина морфологических изменений гоминид от эпохи к эпохе. Мы можем лишь предполагать с большей или меньшей степенью вероятности, чем было вызвано то или иное морфологическое изменение, легшее в основу дальнейшей эволюции. И как раз здесь, используя имеющиеся данные о совр. состоянии палеоантропологии и биологии, я пытаюсь создать мост, который прочно соединил бы прошлое с современностью.
Опять-таки любой биолог скажет, что с течением времени более универсальные формы эволюционной цепи завоёвывают большее географическое пространство в ареале по отношению к предшествующим и постепенно вытесняют последних, как локальные группы, в менее приспособленные для нормальной жизни районы, т. е. в иные экологические ниши, при условии, что не происходит процесс гибридизации (порой даже не посредством скрещивания, а в результате воздействия на организм одних и тех же вирусов) и предыдущая формация не «растворяется» в последующей (что вполне возможно, и часто имеет место). Однако дальнейшая эволюция может приводить (и часто приводит) к тому, что и в этих нишах появляются конкуренты, которые тем или иным образом способствуют либо исчезновению этих популяций, либо превращении их в эндемики — зачастую остатки некогда многочисленных видов, выжившие в каких-то нишах после изменения благоприятных природных условий. Наконец, есть и 3-й путь: под влиянием существенно изменившихся условий обитания — изменение вектора эволюции и получение новых, особенных морфологических черт, отличающих вид как от предкового, так и от родственных.
Человечество насчитывает порядка 8—10 млн. лет, и за этот период полного исчезновения ряда гоминид не произошло, т. е. они нашли себе определённые экологические ниши. Так, например, «дикий человек» (троглодит), как подвид азиатской гориллы, ведёт активный образ жизни в основном в ночное время и населяет те районы Евразии и С. Америки, где присутствие человека минимально или малоощутимо. То же самое можно сказать и по поводу архаических человеческих коллективов — например, андаманцах, негритоссах Малакки и Филиппин, койсанах и пигмеях Африки — архаичных не только образом жизни, но и нек. физиологическими и морфологическими чертами организма: так, у бушменов (койсанские народы) имеются единственные в своём роде особенности — атавистическая постоянная эрекция пениса у мужчин и жировые бёдра у женщин (выполняющие функцию наподобие верблюжьих горбов). Широкого их распространения не наблюдается. Занимаемые ими экологические ниши невелики (для койсанов — в основном пустыни Намиб и Калахари; только австралоидами занят целый континент, на котором ранее, однако, не существовало представителей других рас в силу своей географической изолированности). Легко заметны различия между представителями этих архаичных популяций и более продвинутыми в эволюционном плане монголоидами и негроидами, имеющими с койсанами нек. общие черты. Негроиды распространялись в древности — или населяют сейчас — не только в Африке, но и в Европе, на Кавказе, Б. Востоке, в Ю. Азии и Океании, куда проникли ещё в период нижнего палеолита. Ветвями их стали нек. аборигенные народы Индии, эфиопы, папуасы Новой Гвинеи, меланезийцы, тасманийцы. Это доказывает, что группы негроидов пытались осваивать другие географические регионы с иными климатическими условиями, но не смогли в них удержаться в силу своих физиологических и морфологических особенностей. Т. о. нет никакой основы для того, чтобы отрицать способность того или иного расового типа (вида) к заселению какой-либо географической зоны: необходим лишь стимул и время для адаптации. Однако, появление полиморфизма — расового многообразия, в значительной степени как раз и связано с определённой средой обитания и образом жизни (а также питанием).
Начало формирования культуры на определённом истор. этапе развития привело к нек. выравниванию шансов на существование различных популяций.
Основываясь на теории происхождения вида Homo sapiens от группы завершивших водную стадию приматов-гоминин, можно видеть, что человечество действительно происходит из единого центра. Более древних, чем африканские (ликский и рудольфский из Кении), юаньмоуский и яванский/солоский питекантропы, возрастом более 1 млн. лет, представителей древнейшего человека пока не известно. Более поздними можно считать синантропа, алтайского/денисовского, гейдельбергского людей, и т. н. человека Идалту. Впрочем, не исключено, что к прямым предкам предкам человека относится т. н. чадантроп (с оз. Чад), восходящий к эпохе австралопитеков. В отличие от полицентристов, если принять за основу не архантропный, а более древний период, можно констатировать, что в данном случае они безусловно правы, удревняя в противовес моноцентристам начало формирования рас.
С точки зрения происхождения древнейшего предка человека я стою на позиции моноцентризма, с точки же зрения на происхождение рас — на позиции полицентризма (исходя из законов развития животного мира), объединяя, казалось бы, на первый взгляд, необъединимое. Моя теория антропогенеза объясняет не только происхождение человека, но и его рас, антропологических типов, народов, тесно смыкаясь с этнологией. Кроме того, она позволяет полнее представить эволюционные цепи и животного мира в целом, а также дальнейшие тенденции эволюции. Подтверждение этой теории, существующей без малого 30 лет, нашлось буквально недавно. http://www.pravda-tv.ru/2013/06/13/24747
Вопрос о происхождении совр. человека (неоантропа) в нын. условиях может рассматриваться с 3 позиций. 1-я: путь постепенной эволюционной преемственности формаций (который не мог быть равномерным для всех ныне существующих рас). 2-я: от неб. группы неоантропов, расселившихся по всему Старому и Новому Свету и уничтоживших-де предшествующие формации. 3-я: от подобной же группы, оказавшей вирусно-генетическое влияние на местные формации (опять-таки в пределах всего Земного шара). Совершенно очевидно, что 1-й вариант не мог привести к равномерным изменениям всех существовавших популяций. 2-я теория совершенно нереалистична: неужели ни одна из групп предшествовавших формаций не могла где-либо сохраниться? — Изолированных участков достаточно: Кавказ, Памир, Тибет, Амазония, острова и архипелаги и т. п. И только 3-я версия относительно не противоречит результатам исследований биомолекулярной биологии. Но, однако, принять утверждение биологов, проведших исследование на биомолекулярном уровне о происхождении всего человечества от одного праотца и от одной праматери с упором на формацию неоантропов мы не можем. Волей-неволей мы вынуждены принять единственный вариант того, что данный момент относится ещё к эпохе становления человечества вообще, т. е. становлению первых прямоходящих гоминин, когда появляются первые чопперы (оббитые камни).
В период, когда проблема происхождения Homo sapiens решалась прямолинейно и считалось само собой разумеющимся что совр. человек возник в процессе постепенной эволюции из форм ему подобных (что совершенно неправдоподобно из-за того, что повсеместное смешение или генетическое изменение не могло происходить равномерно по всему Земному шару), считалось, что прародина совр. человека занимала сравнительно неб. территорию, что укладывалось в рамки единой гипотезы узкого моноцентризма. Такая гипотеза сводила на нет роль смешения и обмена техническими достижениями, противоречила она и эволюционным принципам — изолированные группы редко бывают родоначальниками новых видов — убиквистов, т. е. видов, расселённых по всей земной поверхности.
Первым пробил брешь в этих представлениях Ф. Вайденрайх в докладе, сделанном в 1938 г. в Стокгольме на II Международном конгрессе антропологических и этнографических наук. Он насчитал 4 центра возникновения совр. человека по числу выделенных им совр. больших рас — в ЮВА, В. Азии, Африке и Европе. Первый центр послужил зоной формирования австралоидов, второй — монголоидов, третий — негроидов, и последний — европеоидов. Исходными формами для австралоидов он считал яванских питекантропов, для монголоидов — синантропов, для негроидов — африканских неандертальцев и для европеоидов — европейских неандертальцев. В каждом случае генетическая связь древних форм с совр. расами аргументировалась Ф. Вайденрайхом с помощью морфологических сопоставлений: австралоидов сближал с яванскими питекантропами саггитальный валик черепной коробки, который действительно встречается на черепах совр. австралийских аборигенов чаще, чем у представителей остальных рас; монголоиды похожи на синантропа уплощённым лицом и лопатообразными резцами; для европеоидов, как и для европейских неандертальцев, характерны ортогнатный, или мало выступающий вперёд, профиль лицевого скелета и сильно выступающие носовые кости. Только для негроидов такое морфологическое сближение с ископаемыми формами встречается с трудностями, т. к. единственный череп африканского неандертальца с сохранившимся лицевым скелетом из Брокен-Хилла (совр. Кабве) в Родезии (Замбии) отличается крайней ортогнатностью, чем напоминает скорее европеоидов. Но ширина грушевидного отверстия у него, во всяком случае, большая, что свидетельствует о платиринии (широконосости) — существенной особенности негроидной расы. Однако, нам известен с.-афр. тип Мешта-Афалу/Мехта-Афалу (по стоянкам Мешта-эль-Арби и Афалу-бу-Руммель в Алжире), которому также присуща платириния, что может быть свидетельством миграции части предков оного на юг.
В 1962 г. была опубликована общая ревизия новых фактов, свидетельствующих в пользу полицентризма. Автор её, К. Кун, выделил 5 центров, в т. ч. на территории Африки — 2 вместо 1.
Через нек. время моноцентрическая гипотеза была возрождена Я. Я. Рогинским в форме т. н. широкого моноцентризма, который аргументировал положение о том, что трансформация палеоантропа в человека совр. вида могла осуществиться только на достаточно обширной территории, построив всё на морфологической аргументации. Он статистически, по большому числу признаков, отрицал специфическое сходство совр. рас с ископаемыми формами и их генетическое родство, утверждая, что размах изменчивости палеоантропов по многим важным признакам больше, чем у совр. человека, и исключил т. о. из родословной человека все ископаемые формы, кроме палеоантропов группы Схул, в которой им были обнаружены формы, сближенные с совр. расами — монголоидами, негроидами и европеоидами, тем самым фактически отрицая эволюционную теорию. Наконец, центральное положение группы на территории ойкумены позволило ему предположительно очертить ту обширную область, где происходило формирование Homo sapiens: С. Африка (Магриб), В. Средиземноморье (Левант), Кавказ, П., Ср. и Ю. Азия.
Однако в ответ сразу же последовали критические замечания. Г. Ф. Дебец в 1950 г. указал, что характер перехода от эпохи мустье (средний палеолит) к верхнему палеолиту (мадлен), отражённый в археологическом инвентаре, одинаков на тех территориях, которые не входили в очерченную Я. Я. Рогинским зону формирования Homo sapiens. Количество таких фактов, противоречащих гипотезе широкого моноцентризма, постоянно увеличивается. В области археологии было неоднократно доказано непосредственное происхождение многих локальных вариантов верхне-палеолитической культуры от отдельных мустьерских, убедительно продемонстрирована преемственность культуры от среднего палеолита к верхнему на многих узколокальных территориях. Этим постулируется возможность многократного перехода от палеоантропов к человеку разумному — основа основ гипотезы полицентризма. Исходя из всех фактов, можно склониться к выводу о множественности центров появления больших рас Homo sapiens, о формировании разных его рас на основе различных групп ископаемых предков совр. человека.
Если группировать совр. расы по основным морфологическим признакам, то можно выделить 3 или 4 расовых ствола:
1. австралоидный, включающий в себя суданскую расу в Африке (фульбе, в т. ч. водабе), веддоидные народы Индостана и Цейлона, айнов Сахалина, Курил, Хоккайдо, новокаледонцев, австралийских аборигенов, некоторые народы С. и Ю. Америки (калифорнийцы, гуарани, огнеземельцы), и, как подгруппу — европеоидов, делящихся на северную и южную ветви: первая в очень глубокой древности отделилась от австралоидного ствола и заселила север, вторая занимала более южные, присредиземноморские р-ны Европы и Магриба;
2. негроидный, включающий собственно негров, эфиопов, папуасов Новой Гвинеи, негритоссов Австронезии (Андаманы, Малакка, Филиппины и пр.), меланезийцев, тасманийцев (ныне истреблённых);
3. монголоидный, включающий собственно монголоидов, койсанов (бушменов, готтентотов Намибии и хадзапи В. Африки) и
4. америндов (а также т. н. уральцев — угорские народы, селькупы).
Наиболее архаичной формой, имеющей признаки всех больших рас, можно считать пигмеев (не только африканских, но и новогвинейских).
Для австралоидов, как и для негроидов, в своей основе характерны широконосие, тёмная или смуглая пигментация (за исключением с.-европеоидов), чёрные вьющиеся волосы, развитый третичный волосяной покров (усы, борода), долихокранность (длинноголовость) — как у части с.-европеоидов. Европеоиды северной ветви отличаются слабой пигментацией, выступающим узким носом, сильно развитым третичным волосяным покровом (у южных европеоидов нередка сильная волосатость ног, предплечий, груди, часто — брахикранность), долихокранностью.
Негроидам свойственны тёмная пигментация, курчавые волосы, прогнатизм, слабо развитый или отсутствующий третичный волосяной покров, т. н. «овальноголовость».
Монголоидов отличает желтоватая или смуглая кожа, чёрные прямые волосы (для уральской группы характерна светлая пигментация), лопатообразные резцы, эпикантус, уплощённое лицо, слаборазвитый третичный волосяной покров, брахикранность (круглоголовость).
Все эти признаки позволяют более точновыделить прародины больших рас, тем более что в принципе известна прародина их общего предка, а именно — оз. Чад, самой природой разделённое на 3 водоёма, где и оформлялись первичные популяции.
Предположительно, один из древнейших африканских архантропов — рудольфский (оз. Рудольфа (совр. Туркана) в Кении) был непосредственным предком пигмейских племён (имеющих родственников в Новой Гвинее — племя эйпо), населяющих ныне Ц. Африку. Миграция части этих пигмеев за пределы Африки в Азию, по-видимому, положила начало не только новогвинейским пигмеям, а, м. б., и ряду других расовых типов.
В неустановленную пока эпоху в Африке обосновался вид неандертальца, родственного яванскому (известному так же как солоский питекантроп или нгандонгский человек), «исчезнувшему» около 30 тыс. л. н. Не исключено что он является предком суданского типа, к которому в Африке относятся совр. фульбе.
Также в неустановленную эпоху (возможно в верхнем палеолите) в Африку, возможно, попала и ветвь монголоидов (если только койсаны не являются древнейшими африканскими предками монголоидов), часть которых сохранилась в В. Африке (хадзапи), а часть долгое время заселяла Ю. Африку (бушмены и готтентоты). Примечательно, что палео-индейцы Ю. Америки походили на койсанов, но были значительно выше ростом, что говорит о их принадлежности к древнему монголоидному стволу и о том, что уже на руб. 100 тыс. л. н. монголоиды в своей основе давно уже жили в В. Азии. Учитывая относительную близость монголоидов и америндов в морфологическом плане (равно как и значительные различия), а также известное сходство синантропа с гейдельбергским человеком, населявшим Европу, можно выдвинуть осторожное предположение о некоторой предковости «гейдельбергца» с.-америндам. Этот вид мог быть вытеснен из Европы занимавшими приледниковые зоны предками с.-европеоидов, представленных, возможно, т. н. «прогрессивным» питекантропом (в то время как предком ю.-европеоидов мог являться т. н. атлантроп, известный в Магрибе), и сохранил, как и монголоиды, первоначально занимавшие гораздо меньшую территорию обитания, нежели сейчас, смуглую кожу, чёрные прямые волосы и тёмную пигментацию глаз (что характерно и для ю.-европеоидов), и в отличие от них, как у европеоидов, эфиопов, фульбе, части ю.-инд. дравидов — узкий выступающий нос.
Происхождение австралоидной ветви следует связывать, видимо, всё же с ЮВА, откуда эта популяция мигрировала как в Иран и Индостан (ведды, мунда-кола), а непосредственно из ЮВА — в Австралию, Новую Каледонию, в Маньчжурию и Приамурье, и на Японские о-ва, а через них — даже в Америку. На самом древнем этапе гот этой ветви и отделилась европеоидная, часть которой мигрировала на крайний север, в приледниковую зону, и в Европу, приобретя за сотни тыс. и даже млн. лет светлую пигментацию. Вероятно, предками австралоидной ветви были яванские питекантропы, которым в Африке соответствовали ликские (из В. Африки, включавшей Судан как географическую область) — вероятные предки фульбе и водабе.
Всё это говорит о том, что исторический спор о прародине человечества закончен, и прародиной его является, хоть и Африка, но прародиной пра-предка человекообразного вида — Азия. Интересно, что в В. Африке, к востоку от Нила и Большого Африканского Рифта найдено множество останков гоминид — предков человекообразных обезьян, но они известны также в Ю. Азии и ЮВА. Для различных гоминид, кроме человека, Красное море, Персидский залив (в древности — закрытое озеро), а также некогда зелёные Аравия, Ю. Иран, превратившиеся, как и Сахара, в пустыни, стали непреодолимыми преградами для миграций.
*
Сегодня условно можно выделить 8 крупных вида человека, объединённых в один род. Это:
1. африканский/негроидный вид (африканский питекантроп и, в т. ч. юаньмоуский архантроп Ю. Китая (Юньнань), 1,7 млн. л. н.);
2. айнский тип (гейдельбергский человек, 430 тыс. л. н.);
3. афро-европейский/фульбоидный тип (человека Идалту, 170 тыс. л. н.);
4. южно-европейский/средиземноморский вид (схулский неандерталец);
5. северо-европейский вид (кроманьонец);
6. алтайский/америндский вид (алтайский/денисовский питекантроп/палеоантроп);
7. австронезийский/австралоидно-веддоидный вид (яванский/солоский питекантроп);
8. азиатский/монголоидный вид (синантроп, в т. ч. прото-монголоидные койсаны Ю. и Ц. Африки).
При этом африканский вид является не монолитным, а состоит, по меньшей мере, из 3—4 видов. Между ними различия больше, чем между всеми остальными, вместе взятыми.
Ок. 2 млн. л. н. на Яву пришёл яванский/солоский архантроп, носитель гаплогруппы С. Ок. 1—0,5 млн. л. н. от него отделился гейдельбергский архантроп (гаплотипы С1а1 и С1а2).
Ок. 1,7 млн. л. н. от африканского питекантропа, носителя гаплогрупп А и В, отделился юаньмоуский архантроп (гаплогруппа D).
Ок. 1—0,5 млн. л. н. в С. Китае появился синантроп (гаплогруппы О и N).
Примерно 700—500 тыс. л. н., от, условно «общего ствола» отделился алтайский/денисовский архантроп (гаплогруппа Q). Это значит, что между любым человеком, живущим в центре России, и человеком, живущим в ЮВА генетическое расстояние такое, которое сформировалось за 1—1,4 млн. лет.
Далее от «общего ствола» отделился схулский/восточный неандерталец (гаплогруппы F, H, G, K, Т, L, M, S, J, I и пр.), или средиземноморский вид. Это, по разным данным, состоялось от 400 до 200 тыс. л. н. Т. е. между нами и нек. средиземноморцами генетическое расстояние может достигать 800 тыс. лет.
Затем появился пре-сапиенс, кроманьонец (гаплогруппы P, R), с.-европейский вид.
И последним от «общего ствола» — примерно 170 тыс. л. н. (что совпадает с поздней планкой денисовца) отделился один из африканских видов эфиопский человек Идалту (гаплогруппа Е), предок фульбе. Это вид уже жил в Азии, точнее, на Б. Востоке, откуда мигрировал как далее в Евразию (!), где уже были свои виды человека, так и в Африку.
В то время как основная масса представителей гаплогруппы С имеет папуасо-веддо-австралоидные расовые типы, ею одновременно обладают ц.-аз. широколицые монголоиды и палео-азиаты, проникшие даже на крайний с.-з. С.-Амер. континента. Возникает закономерный вопрос: каким образом и когда нын. типично-монголоидные народы Ц. и отчасти В. Азии приобрели прото-австралоидную гаплогруппу С? Их предком не может быть ни алтайский/денисовский архантроп/палеоантроп (Хомо алтаенсис), пра-предок америндов гаплогруппы Q, ок. 700 тыс. л. н. отколовшийся от гейдельбергского архантропа/палеоантропа (предка прото-айнской гаплогруппы С1а2, как вид, обосновавшегося в Европе), и существовавший там ещё 40 тыс. л. н. Им не может быть ни с.-кит. синантроп гаплогруппы О, ни юаньмоуский архантроп из Ю. Китая (Юньнань), от 1,7 млн. л. н., носитель гаплогруппы D, прямой предок как основной массы айнов, так и в т. ч. островных негроидных андаманцев, оторванных от континента — но зато это могут быть все австралоидные народы, восходящие к яванскому/солоскому питекатропу, жившему 2,0—1,6 млн. л. н. — 200 тыс. л. н. Следовательно, у нас имеется неск. типичных ископаемых претендентов на роль носителя айно-папуасской гаплогруппы С1 (с кластерами С1а1, С1а2, C1b) — Костёнки-14, Сунгирь-2, Арене Кандиде. Носителем фенотипа (расового типа) в Ц. и В. Азии стал с.-кит. синантроп, живший ок. 770±80 тыс. л. н., предок всех монголоидов, носитель гаплогруппы О, принявший основное участие в метисации с западными пришельцами.

Волны прото-гаплогрупп С и D (яванский и юаньмоуский архантропы/палеоантропы) мигрировали из Азии через Ц. Азию на Кавказ и в Европу, а также в Ю. и С. Китай, где фиксируются ископаемые виды алтайского/денисовского архантропа/палеоантропа и синантропа, положив т. о. начало 4-м расам — айно-веддо-австралоидной, негритосско-папуасской, америндской и монголоидной. Не исключён также вариант проникновения юаньмоуского питекантропа из Ю. Китая в Ц. Азию, где могло произойти смешение с синантропом (народ пуми, у которого частота D1a2a (P47) составляет 70%). Такая миграция могла идти только к истокам по руслу великих ю.-кит. рек, Салуина и Меконга, смыкающихся с истоками великих в.-кит. рек — Хуан-хэ и Ян-цзы. Следовательно, где-то в округе их истоков должна находиться прародина монголо-тибетских народов, которые, однако, выходят на арену истории довольно поздно — к руб. н. э., когда прото-монгольские племена сяньби, выдвинувшиеся из В. Маньчжурии, захватили ареал тюркоязычных народов хунну. Однако, нам известно, что маньчжуры и тибетцы восходят к одним истокам, лежащим как раз в истоках великих ю.-кит. и в.-кит. рек.
Всё это помогает понять, что все «расчёты» т. н. ДНК-генеалогии притянуты за уши и нисколько не отвечают известной палеоантропологической картине.
Глава 5 Формации и виды Homo sapiens. Полиморфизм
Каковы основные побудительные причины, приведшие к появлению 3—4 условных человеческих формаций?
Первое. На этапе перехода гоминид к водно-собирательному образу жизни начались оформление локомоции кисти и переход к выпрямлению позвоночника, и, как следствие — прямохождению, сопровождавшиеся редуцированием (уменьшением), а позже — и исчезновением волосяного покрова. Окончание водной стадии и появление гоминин в новых экологических нишах существенно сузило экологические ниши австралопитеков — предков афр. обезьян, которым, не исключено, принадлежала и Олдувайская культура с её чопперами. По всем морфологическим признакам и последним данным молекулярной биологии, наиболее архаичной, аморфной в расовом плане формой человека могут считаться совр. ц.-афр. пигмеи (ака, бамбути и пр.), в соответствии с законами животного мира, после возникновения древнейшей популяции начавшие деление на отдельные группы, от которых, в свою очередь, произошли другие формы, заложившие основу ещё на этом, древнейшем, этапе, разделения человечества на расовые стволы, имевшие общие морфологические признаки — форму черепа, волос, строения зубов, пигментацию. Так, например, койсаны — одни из аборигенов Африки наряду с пигмеями, имеют значительное морфологическое сходство с монголоидами (жёлтый цвет кожи, уплощённое лицо, эпикантус, лопатообразные резцы). Койсаны и негритоссы (Андаманские о-ва, Малакка, Филиппины) являются предками двух больших расовых ветвей — монголоидной и негроидной, которые возникли путём перехода в новое качество — отделением и разделением популяций от основной предковой массы в соответствии с классическими законами животного мира. Т. о. можно говорить о начале процесса появления рас, на самом древнем уровне — уровне предков ныне существующих примитивных низкорослых народов мира экваториального пояса. Эти народы ещё не были готовы к заселению арктических широт, которое началось, возможно, уже 2 млн. л. н.
Думается, что «перво-люди» изначально не были курчавы (как не бывает курчавых гоминид), и нек. группы стали таковыми в процессе расселения во влажной экваториально-тропической зоне: в их число, кроме негроидных представителей, входят и койсаны — предковая монголоидная популяция. Наиболее архаичные представители австралоидного ствола имеют вьющиеся волосы — ведды, населяющие Цейлон, и родственные им народности Ю. Индостана. Т. о. возможно предполагать, что они стоят ближе к «перво-людям» (гомининам), чем негритосы и койсаны.
Второе, Олдувайская эпоха кончилась с переходом к Шелльской, т. е. тогда, когда произошло кардинальное изменение конституции гоминин в сторону массивности, сопровождавшееся резким увеличением мозга и появлением зачатков речи, стимулируемое переходом к охоте и увеличением потребления белковой (животной) пищи. Одновременно происходило изменение нижней челюсти и жевательной мускулатуры. Переход к охоте стимулировал как увеличение мозга, так и общее изменение морфологии. Появилась новая формация — Homo erectus (человек прямоходящий) или архантропы (древнейшие люди), которые, отличались значительно большей массивностью и значительно большим ростом, были более умными и развитыми, а также подготовленными к различным природным условиям, и, соответственно, более конкурентоспособными. Их ископаемые останки встречаются уже в недоступной малорослым теплолюбивым экваториальным популяциям северной полосе, в т. ч. в Европе, в непосредственной близости от ледниковой зоны.
Третье. Начавшаяся Мустьерская эпоха отличалась от предыдущей главным образом появлением собственно культуры, окончательным оформлением морфологии тела и качественной перестройкой морфологии мозга, выразившейся в увеличении полушарий и редуцировании мозжечка. Палеоантропы уже владели в известной степени речью монологового или отрывочного типа, умели строить жильё и шили одежду, т. е. их образ жизни уже складывался под воздействием не только природных, но и искусственных факторов.
Этот этап характеризуется, в первую очередь, тем, что эволюция Homo стала приобретать новые формы морфологического развития, которые вырабатывались под влиянием условий, уже в значительной степени искусственных, не характерных для животного мира, поскольку ещё на этапе архантропов началось формирование древнейших форм человеческой культуры, развившейся в эпоху палеоантропов. Он характеризуется также тем, что здесь природа подвела Homo к своеобразному физическому пику, когда прекрасно приспособившийся к различным, в т. ч. и крайне суровым средам обитания, человек достиг своего физического предела: зачастую его скелет, объём черепной коробки, мышечная структура превышают показатели, характерные для последующего, более грацильного, но и более организованного неоантропа.
На каждом качественно-новом этапе существовали свои культурные пороги, через которые не всегда легко было переступать более примитивным популяциям.
Четвёртое. Условия развития палеоантропов привели к новой тенденции — усилению социального фактора, что отразилось на дальнейшем формировании морфологии головного мозга, выразившейся в увеличении лобных долей, функции которого значительно усложнились. Черепная коробка приняла совр. вид. Зачастую, под влиянием искусственных условий, выражавшихся в новом образе жизни, начала проявляться грацилизация организма. Для этого же периода характерен отказ от каннибализма.
Последний этап — эпоха неоантропов, начавшаяся ок. 200—150 тыс. л. н., характеризуется появлением более утончённых, значительно более развитых и приспособленных к любым природным условиям морфологических черт. В первую очередь, это относится к строению головного мозга, и, соответственно, черепной коробки. Следует отметить более грацильное сложение неоантропов, что, по-видимому, является признаком недостатка белковой пищи, и, соответственно, более южного происхождения. Как ныне принято считать — их формирование происходило на базе всех популяций с участием генома наиболее развитого вида (в той или иной степени — авт.) — а именно северного, через стадию т. н. «прогрессивных» палеоантропов. Нек. популяции, в т. ч. негроидные, австралоидные, монголоидные, получившие наибольший процент данного гипотетического генома, отличаются наличием характерных признаков большей морфологической прогрессивности по сравнению с классическими типами. К ним, например, относятся эфиопы, з.-афр. фульбе, нек. ю.-инд. народы и отдельные группы монголоидов.
Т. о. мы можем наблюдать определённые этапы эволюции человечества и качественные показатели каждого периода, соответствующие уровню культуры и т. о. уровню морфологического развития головного мозга, из чего следует заключение о различных уровнях возможностей дальнейшего развития ныне существующих расовых типов, что, впрочем, не следует путать с понятием расовой неполноценности, утрирующим этот вопрос, поскольку у каждой расы изначально были разные условия для развития.
В то же время нельзя говорить и о завершённости эволюционного процесса, ведь мы живём в мире, где представлены все расовые группы, сформировавшиеся на базе 4-х условных формаций человеческого общества. К тому же в обществе происходит всё более хаотическое смешение расовых типов, что необратимо должно привести к качественно-новому скачку эволюции, основанному на несравненно большем разнообразии генного потенциала. Самая большая проблема, которая при этом возникает — невозможность прогноза инбридинга, чреватыми тяжёлыми наследственными недугами, способными уничтожить значительную часть населения, оставив наиболее жизнеспособных, иммунно-защищённых индивидов, что, впрочем, не означает того, что антропологически они будут однородны, если будет происходить процесс рецессивного выщепления (т. е. это будут наиболее морфологически-совершенные типы): расовое деление (полиморфизм) сохранится. Скорее всего, это будут те, у кого наиболее разнообразен генетический набор. Именно общество этой формации будет носить определение Homo sapientissimus.
Собственно говоря, все этапы перехода на новые формации не имеют чётких границ, поскольку эти процессы происходили в течение сотен тыс. лет. Так, когда ставится вопрос о языковом развитии австралопитековой формации, можно сказать, что для неё речь в собственном смысле этого слова ещё не существовала, хотя у австралопитеков уже имелись нервные центры Брока и Вернике, способствующие возникновению речи. Произошло лишь качественное усложнение системы коммуникации, основанное на гораздо более мощной, чем у обезьян, вокально-информативной системе. Соответственно, увеличение команд и сообщений находило выражение в разных гортанных, носовых, щёлкающих звуках, очень характерных и сейчас для примитивных народов (в частности, пигмеев, койсанов).
Несмотря на то, что в известной мере все человеческие популяции, трансформировавшиеся в т. н. неоантропов, на первый взгляд, как будто застыли на нын. морфофизиологическом уровне, это ещё не означает, что они не продолжают процесс внутреннего самосовершенствования. Впитывая гены и культуру разных формаций, они морфологически совершенствуют, в первую очередь, свой мозг, который особенно на последней стадии развития человечества стал сближаться по объёму и морфологии у различных популяций. Происходило и нек. внутривидовое совершенствование конституции, давшее нын. результаты. То, что обычно называется племенами — у примитивных народов не есть племена в прямом смысле этого слова — это, скорее, родовые общины во главе с вожаком (вождём), занимающие определённые экологические ниши, наряду с прочими представителями животного мира, в т. ч. и обезьяньими группами, в отличие от которых венчают пищевую пирамиду, и т. о. неразрывно связаны с окружающей их природой. Так, к примеру, пигмеи не имеют своего собственного языка для внутриплеменного общения, и для общения с соседними негроидными народами пользуются их языком.
Известно, что вид эволюционирует быстрее всего, когда он разбит на популяции, между которыми до какой-то степени осуществляется обмен генами. Непостоянные контакты между популяциями могли поддерживать на определённом уровне обмен генов, способствовавший прогрессивной эволюции.
Говоря о происхождении человеческих формаций, проявлении полиморфизма, миграциях и расселении рас в определённых ареалах, следует остановиться на самом процессе смены формаций, на самом процессе эволюции.
Мутации довольно часто бывают рецессивными, т. е. до поры никаких изменений у новорождённых не наблюдается. Дети рождаются такими же, как и их родители. Но при изменении климата, ареала расселения «вдруг» начинают появляться новые виды. Это значит, что стали действовать скрытые (рецессивные) мутации.
В изолированных друг от друга популяциях половые связи возможны только внутри группы между родственниками — явление, которое биологи называют инбридингом. Инбридинг приводит к тому, что линии очищаются от примесей и становятся пригодными к гибридизации, т. е. при скрещивании с другими линиями дают новое, более способное к жизни, потомство.
Знаменитый скульптор-антрополог М. М. Герасимов, анализируя погребения эпохи Мустье из пещер горы Кармел в Палестине, отмечал: «Создаётся впечатление, что в скелетах мустьерцев из Палестины зафиксированы эволюционные этапы трансформации неандертальцев в новый вид человека. И этому можно было бы поверить, если бы скелеты, найденные в Палестине, были распределены в культурных горизонтах следующим образом: примитивные — в нижних слоях мустье, смешанные формы — в средних слоях мустье и в слоях финального мустье — люди типа Схул IV и Схул V. В данном случае это было не так. И наиболее примитивные люди (Схул IX и Табун I), равно как и Табун II, Схул IV и Схул V, жили примерно в одно и то же время, на протяжении относительно короткого срока культуры леваллуазского мустье».
Почему обязательно вымирают предковые формы? — Если принять гипотезу о том, что изменение обуславливается мутацией, то появление новых типов — это прежде всего результат активизации мутации, т. е. ответ организма на изменение окружающей природной среды. Старые организмы вымирают потому, что не могут переносить новых условий, а новые живут потому что само появление их — результат приспособления организма к этим условиям. Если природные условия резко менялись, то и ответная реакция организма была быстрой, т. е. активизировались рецессивные мутации.
Сосуществование палеоантропов и людей, близких к совр. типу в пещере Кармел и есть свидетельство того, что мутации настолько быстро действовали, что предки и потомки какое-то время жили вместе в одной пещере.
Если группа была длительное время разделена какими-то географ. барьерами на 2 части, то у каждой из них появляются настолько существенные отличия в строении организма и поведении, что потом, когда этот барьер исчезает, они уже не могут скрещиваться и давать плодовитое потомство. Эти 2 группы становятся отдельными видами.
У человека на уровне рас никакого зримого генетического расхождения не наблюдается, поскольку люди разных рас происходят от одних предков. Изоляция отдельных групп человечества хоть и привела к появлению рас, но не была настолько длительной, чтобы привести к обособлению их в разные виды.
Т. о. возникли архантропы, палеоантропы и, наконец, неоантропы. И всегда разные формации нек. время сосуществовали друг с другом. Поскольку перестройка морфологии — процесс длительный, и уж коль скоро мы принимаем наличие неандертальской фазы в эволюции человека, то должны сделать вывод о формировании отличительных морфологических особенностей неоантропов в недрах групп палеоантропов, и сосуществовании палеоантропа и неоантропа (а быть может, и архантропа) на определённом отрезке времени теоретически неизбежно. Сосуществование более примитивных и более прогрессивных морфологических форм составляло характерную черту эволюции гоминид на всех этапах их истории. Т. о. процесс становления Homo sapiens был довольно длительным, а, следовательно, и сложным.
Исследователь синантропа Ф. Вайденрайх считал наиболее показательным отличием совр. человека от палеоантропа совершенный по своей структуре мозг — с более развитыми полушариями, увеличенный в высоту, с редуцированным затылочным отделом. Я. Я. Рогинский, выдвигая гипотезу о факторах формирования человека совр. вида, использовал широко известные в клинике нервных болезней наблюдения над субъектами, у которых повреждены лобные доли мозга (т. н. лоботомия — авт.): у них резко тормозятся или совсем пропадают социальные инстинкты, буйный нрав делает их опасными для окружающих. Т. о. лобные доли мозга — средоточие не только высших мыслительных, но и социальных функций. Этот вывод был сопоставлен с установленным на эндокранах фактом разрастания лобных долей мозга у совр. человека по сравнению с палеоантропом. Рогинский указывал на огромную общественную силу и жизнеспособность тех коллективов, у которых число социальных особей было наибольшим. Разрастание лобных долей расширяло сферу высшего ассоциативного мышления, а с ним способствовало усложнению общественной жизни, разнообразию трудовой деятельности, вызывало дальнейшую эволюцию тела, физиологических навыков, моторных функций.
Совершенно очевидно и естественно, что на протяжении млн.-в лет предки людей много раз делились на ветви, давая начало новым популяциям — малым расам, расовым типам, племенам. Уже с наступлением архитектурной эпохи мы начинаем сталкиваться с большим разнообразием, когда уже проявились черты полиморфизма, свойственные той или иной большой расе.
Считать полиморфизм совр. человечества явлением позднейшей стадии неверно. Являясь убиквистом, человеку, чтобы заселить разнообразные географические среды, приходилось расширять и интенсифицировать адаптивные реакции, что и привело к дифференциации форм и сочетаний изменчивости, и т. о. к полиморфизму, что и закреплялось отбором. Для этого был необходим значительный временной период, и начало было положено, по-видимому, ещё в начале нижнего палеолита при первых шагах заселения ойкумены. Полное её освоение представляет собой внешний стимул для возникновения многообразия изменчивости, а сам полиморфизм — уже нек. итог эволюции.
Нельзя не только отрицать, но и необходимо согласиться с фактом того, что локальные варианты мегатипов человека на различных этапах своей истории, испытывали генетическое влияние соседей, и, что само собой разумеется, культурное влияние, менявшее отчасти образ жизни и т. о. служившее возникновению полиморфизма.
Вопрос о возможности группировки больших рас в более общие и крупные категории восходит ещё к работам Томаса Гексли, предполагавшего ещё в 1870 г. делить всё человечество на 2 большие группы. Позднее Я. Я. Рогинский привёл достаточно убедительные данные в пользу разной возрастной динамики расовых признаков 3 больших рас: у европеоидов и негроидов выраженность расовых признаков усиливается с возрастом, тогда как у монголоидов она ослабевает. Но что это нам даёт на практике — непонятно.
При обсуждении трудностей, с которыми сталкивается гипотеза полицентризма, часто фигурирует противоречие полицентристского подхода идее видового единства человечества. Однако, мы сталкиваемся в данном случае с простым недоразумением, проистекающим из неучёта того обстоятельства, что формирование видового комплекса Homo sapiens и расовых комплексов охватывало разные комбинации признаков, и поэтому могло идти параллельно на протяжении длительного отрезка времени. Территориальные вариации в строении вертикального и горизонтального профиля лицевого скелета, микро-рельефе внутренней поверхности резцов, других признаков, не могли помешать формированию видового комплекса, выразившегося в перестройке мозга, повышении высоты черепа, редукции черепного, и, в первую очередь, надбровного и затылочного рельефа, образовании вполне совр. по своим очертаниям подбородочного выступа. Коль скоро действовала тенденция образования совр. вида человека, она охватывала многие популяции, различавшиеся во многих признаках, и подчиняла себе их эволюцию, придавая ей направленный характер перестройки морфологии на совр. уровень. Впервые данную мысль высказал член-корр. АН СССР В. П. Алексеев. Однако не могу согласиться с ним по вопросу формирования первичных очагов расообразования, относимых им ко времени не ранее конца нижнего палеолита (частично), поскольку из схемы расообразования т. о. выводятся более древние группы человеческих предков, хотя многие исследователи включают в их число даже Homo habilis.
Расселение человечества по земной поверхности не было само по себе единственной причиной расообразования. Дифференциация происходила и вследствие установления дополнительных генетических барьеров, частично отражавших географы. рубежи, а частично возникших из-за увеличения расстояний между разделившимися группами. В ряде случаев сходство даже по ведущим признакам могло-де носить конвергентный характер, выражая результат параллельного формообразования, вследствие сходных мутаций (я не согласен с этим утверждением, поскольку такие варианты маловероятны, им пока ничем не подтверждены — авт.). Иная тенденция возникла в процессе этнокультурной консолидации: сама интеграция популяции приводила к распространению определённых генетических комплексов на обширной территории.
Очевидно, что раса — межгрупповое понятие, поскольку в ней налицо отличие популяций от других родственных или географически близких популяций по внутригрупповым коэффициентам. Когда же они сближаются с межгрупповыми — популяция не является гомогенной и имеет сложное происхождение из неск. групп, которые ещё сохранили свою специфику.
Если в гомогенных популяциях коэффициенты внутригрупповой корреляции между одноименными признаками одинаковы по направлению и близки по абсолютной величине (в них находят выражение одинаковые в целом для человечества законы вариаций изменчивости морфологических особенностей), то коэффициенты межгрупповой корреляции тех же признаков часто очень значительно отличаются по своей абсолютной величине от внутригрупповых, а иногда отличаются от них и по направлению.
Смешанное происхождение группы вероятно в том случае, если распад внутригрупповых связей (выражающийся в понижении коэффициента корреляции, а тем более превращении его из положительного в отрицательный) имеет место не в 2, а в неск. признаках (подобный анализ проводил в 1930-х гг. советский антрополог А. И. Ярхо при рассмотрении происхождения многих народов Ср. Азии и Кавказа).
Популяционный подход к расе объясняет тенденции эволюционного развития этносов, поскольку раса представляет собой не индивидуально-типологическое, а групповое понятие, т. е. раса охватывает группу популяций. Изменение же расовых признаков происходит не только в результате внешних экзогенных влияний, например, смешения, но и внутренних популяционных процессов — направленного изменения признаков во времени (включая т. н. «дрейф генов» и т. п.).
Другой вопрос: комплексы тесно связанных между собой морфо-физиологических признаков, выражающих общие тенденции интеграции человеческого организма, лежат в основе выделения конституциональных габитусов и не имеют, как правило, закономерного географ. распространения, варьируют более или менее мозаично. Независимые или слабо зависимые друг от друга в морфо-физиологическом отношении признаки, наоборот, закономерно варьирующие географически, в комплексах образуют расы. В этом их кардинальное различие. А именно раса тесно связана с историей и географией человечества.
Аллопатрический процесс (дифференциация популяций только на географ. основе) — основа видообразования и формирования внутри вида низших таксономических категорий, в отличие от симпатрического (дифференциация популяций на одной и той же территории лишь вследствие приспособления к разным условиям среды).
Существует ряд экологических правил Д. Аллена, К. Бергмана, К. Глогера, А. Томсона и Л. Бакстона. 1-е трактует связь с климатом пропорций тела: в холодном климате укорачиваются конечности и появляется более плотное сложение. 2-е наблюдает зависимость между температурой среды и размерами тела: на севере и на юге, в р-нах сурового климата, размеры тела больше, чем в тропической зоне, 3-е устанавливает интенсивность окраски (пигментации) в зависимости от географической широты местности: чем ближе к тропикам, тем окраска интенсивнее. Последнее, 4-е, выражает зависимость ширины носа от климатических показателей: максимальная ширина характеризует группы человечества, расселённые в тропическом поясе, минимальные величины характерны для арктическо-антарктических зон. С этими правилами можно было бы не согласиться или поспорить: в любом из них есть несоответствия или исключения, однако, мне кажется — дело в сложных путях истор. реализации данной дифференциации. Однако, это как раз доказывает длительность процесса расообразования.
Совершенно очевидно, что любой антропологический тип складывается из популяций, имеющих близкую географию. Расселение неоантропов из одного р-на по Зе6мному шару не могло дать в короткий срок подобной пестроты морфо-физиологической дифференциации без, по меньшей мере, смешения с местными, аборигенными популяциями. Тем более мы не можем отрицать широты определяемой морфологией тенденции развития в конце неандертальского (мустьерского) периода (основанной в т. ч. и на культурных контактах), приведшей к возникновению новой формации — неоантропа.
В самом деле, трудно объяснить появление специфических морфологических черт, исходя из географии только для неоантропов, в то время когда известно, что у предшествовавших им формаций также имелись характерные черты, несмотря на неск. иную географию. Многое, конечно, можно объяснить приспособлением к определённым географ. условиям: ширина носа, курчавость, укороченные конечности, пигментация и многое другое. Но как объяснить уплощение лица монголоидов, прогнатизм и вытянутость черепа негроидов, миниатюрность нек. африканских, северных и океанийских народов? Ведь данные группы живут в тесном соседстве с другими, часто аборигенными, но резко отличающимися от них по ряду морфо-физиологических особенностей. Можно, конечно, чисто гипотетически предполагать, что например, северные лопари (саамы), угроязычные манси и ханты, самодийские селькупы некогда входили в круг «пигмейских» народов экваториального пояса (хотя трое последних относятся к северной, смешанной с с.-европеоидами, ветви монголоидной расы). Однако, во-первых, генетика говорит совсем о другом, а, во-вторых, между афр. койсанами (бушменами, готтентотами, ц.-афр. хадзапи) и пигмеями, не говоря уже о негритоссах Андаманских о-вов, Малакки и Филиппин (аэта, семанги), существует значительное различие, хотя, без сомнения, эти народы — одни из первых аборигенов Африки. Карликовость — генетическая особенность, закреплённая у данных популяций и перешедшая к ним от предков.
Прежде чем перейти к рассмотрению видового и расового деления, следует заметить, что не все экологические правила абсолютно истинны. В частности, правило К. Глогера, как будто, противоречит наблюдаемой ныне картине, а именно — обитающие в экваториальной тропической зоне пигмеи и койсаны (бушмены) не имеют интенсивной пигментации — первые со светло-коричневой кожей, вторые — вообще желтокожи. Кроме того, у бушменов наблюдается уплощение лица, эпикантус, характерные для монголоидов, причём это не конвергентные признаки — у них в т. ч. лопатообразные резцы, как у классических монголоидов. Следовательно, речь не может идти о недавно приобретённой особенности, поскольку и те, и другие являются древнейшими жителями Африканского континента. При сравнении пигментации с.-монголоид. народов (эскимосов, чукчей, коряков и пр.) сразу же бросается в глаза всё тот же ряд сходных признаков — уплощённое лицо, эпикантус, лопатообразные резцы. Но нам известно, что самые древние из них населяют Север не более 3 тыс. лет.
Другой пример: айны (в значительной степени относящиеся к Y-хромосомной ДНК гаплогруппе D1, хотя с частотой ок. 15% также встречается гаплогруппа C3, что является весьма спорным моментом, т. к. за именно японскую C3a (M93) выдают айнскую гаплогруппу С1а1 — М.Р.), как обычно считается, являющиеся северной ветвью австралоидов (у аборигенов гаплогруппа D не встречается, а характерна С4; у расово родственных веддоидных народов Индии и ЮВА — С5, у папуасов и меланезийцев, частично — микронезийцев и полинезийцев, как вторичного субстрата — С2), зап. предки которых в верхнем палеолите населяли даже Север В. Европы (родственный кластер (С1а2) Y-хромосомной ДНК гаплогруппы С, кластер которой — С1а1 — отмечен в Японии), имеют достаточно светлую (смуглую) кожу, но, в результате 2–тыс.-летнего смешения (подавившего исходный австралоидный гаплотип С1а1 японской гаплогруппой D1b), сближаются с монголоидами. Нек. антропологи считают их древней. населением Японского архипелага (а также Сахалина и Курил), заселённого ими более 100 тыс. л. н. Следовательно, посветление (неполное) произошло и было закреплено на генетическом уровне значительно позже эпохи переселения из экваториального пояса (с о. Ява, где фиксируется предок австралоидов — яванский питекантроп, живший 2,0—1,6 млн. л. н. — 200 тыс. л. н.).
Не менее характерен пример с кроманьоидным населением Канарского архипелага, относимого к антропологическому типу «Мешта/Мехта-Афалу», характеризующему часть населения С. Африки (в частности, среди совр. берберов — кабилов и риффов), и присущего части древ. населения З. Европы (в т. ч. Пиренеев). Однако, у с.-африканцев, относящихся к данному типу, заметно выше носовой показатель и присуща платириния (широконосие), что нек. трактуется как негроидная примесь. Однако, «канарцы», при всей своей массивности, отличаются от древней. формы европеоидной расы значительно большей грацильностью. Для данного антропологического типа характерен высокий рост, долихоцефалия, широкое лицо с низкими орбитами, а также светлая пигментация (которая могла появиться лишь в специфических северных условиях), светло-каштановые и белокурые волосы (иногда с рыжеватым оттенком), светлый цвет глаз (чаще — зелёный, реже — голубой и серый). Тем не менее, данный тип не равнозначен нордическому типу, населявшему достаточно поздно северную часть о. Гран-Канария. Этим я хочу сказать, что исходный тип, восходящий к обще-с.-европ. типу, должен был претерпеть ряд морфологических изменений, и, в первую очередь, приобрести светлую пигментацию, в течение достаточно долгого периода, что в условиях С. Африки было невозможно. Следовательно, этот тип мы можем рассматривать только как пришлый сюда не ранее 3 тыс. до н. э.
Наличие же негроидов в Европе в верхнем палеолите-мезолите можно объяснить их притоком сюда одной из волн древ. миграции в период межледниковья из б.-вост. и с.-афр. регионов, где они были представлены достаточно широко.
*
Итак, как мы можем практически представить процесс возникновения полиморфизма — многообразия человеческого сообщества? Учитывая характер эволюции, можно утверждать, что расселение человечества ещё на архи-древнем этапе его истории послужило стимулом формирования локальных расовых групп. В среде древнейших людей — архантропов — возникли Шелльская и Ашельская каменные культуры.
Как-то странно выбивается из среды австралопитеков находка на оз. Чад т. н. чадантропа, останки которого по абсолютному возрасту оказались значительно древнее ю.-афр. австралопитеков (более 2,5—3 млн. лет): если до того основные их находки группировались в Ю. и В. Африке, то это — почти единственная находка подобного рода, исключая азиатских гигантопитеков и афро-азиатских мегантропов, сделанная за пределами этой зоны Африки. Не исключено, что в этом случае речь действительно идёт о собственно человеческом предке.
Как показали результаты биомолекулярных исследований, древнейшие люди были похожи на совр. пигмеев (точнее, были прямыми предками пигмеев), а наиболее архаичными среди последних считают племя ака Ц. Африки, где они живут с самого момента своего здесь появления.
В то же время в Ю. и В. Африке живёт группа койсанских племён — бушменов и готтентотов (ещё одна группа — хадзапи — населяет Ц. Африку) — древнейшей монголоидной ветви. Но не надо забывать, что в период между 2,7—2,2 млн. л. н. в Ю. Африке отмечается австралопитековая формация прямоходящих грацильных плезиантропов (африканских австралопитеков), зубная система которых близка синантропам Ц. Азии — прямым предкам монголоидов. Возникает соблазн возводить койсанов к плезиантропам, и считать их прародителями одной из древнейших рас, превалирующей сейчас в Азии, куда она мигрировала из Африки, если только признать плезиантропа предком архантропов, а не нын. шимпанзе, с которыми у них очень много общего. Проблема в том, что в т. сл. надо признать плезиантропа — предковой человеческой формацией, причём монголоидной расы, а нек. др. виды — предками негроидной и австралоидной рас. Это пока не приходит в противоречие с тем, что наиболее древ. прото-расой были пигмеи, возможные потомки чадантропа, но в т. сл. необходимо доказать их непосредственную генетическую преемственность, и признать, что общий предок человека оформился в Ц. Африке, в р-не палео-озера Чад.
Еще в сер. 20 в. оз. Чад входило в пятерку крупнейших водоемов Африки. Однако с тех пор его площадь резко уменьшилась. Если в 1960-е гг. оно занимало территорию в 26 тыс. кв. км, то к 2007 г. этот показатель уменьшился до 1350 кв. км. Однако все эти цифры не идут ни в какое сравнение с тем, что из себя представляло оз. Чад в относительно недалеком прошлом. Всего 7 тыс. л. н. оно по своему размеру было сопоставимо с Каспийским морем. Его акватория простиралась примерно на 400 тыс. кв. км. Для обозначения этого исчезнувшего водоема ученые используют термин «Мега-Чад» или «Палео-Чад».
Такая трактовка требует объяснения, как в т. сл. предок человека, осваивавший водную среду, попал в Ю. и В. Африку, каковы были побудительные причины к таким миграциям. Она же требует пояснения, почему сухопутный вид пра-человека обосновался именно в Ц. Африке, а его водный вид населил океан, а также встречается в таких закрытых водоёмах как Каспий и Байкал.

Абрис древнего оз. Мега-Чад на фоне нын. озера Чад и границ нын. афр. стран.
На сегодняшний день возраст моджокертского питекантропа, найденного на о. Ява, оценивается в 1,7—0,2 млн. лет. Для найденного в Кении ликского питекантропа наиболее древ. датировка — 1,5 млн. лет. Более поздние его останки встречаются также и в Танзании. Напомню, к западу от Кении и Танзании находится большое оз. Виктория (быв. Рудольфа). Для этого вида отмечаются близкое сходство с яванским (моджокертским) видом, включая аналогичный (или неск. больший) объём мозга (850—1100 куб. см; для сравнения, у яванского — 850—1000 куб. см), что было бы неудивительно, если предполагать миграцию в В. Африку более древнего «яванца». Не исключено, что потомками ликского питекантропа были совр. «суданцы» (фульбе) — ветвь австралоидной расы (прото-австралоиды), предком которой, видимо, и был яванский питекантроп, у которого имеется определённое сходство с австралоидами. Однако, скорее, всё же австралоиды — такие же древ. выходцы из Африки, как и монголоиды. Вид ликского питекантропа «исчезает» только на руб. 300 тыс. л. н., «уступая место» какому-то другому, преемствующему ему виду.
Высыхание оз. Чад в современности
В Кении также были найдены ископаемые останки гоминин возрастом ок. 1,9 млн. лет (по нек. оценкам, до 2,5 млн. лет), рядом антропологов выделяемых в вид рудольфского питекантропа, отличающегося от 2 первых неск. меньшим объёмом мозга (770—800 куб. см, у женских особей — на уровне обезьян и автралопитеков — 500 куб. см), но в то же время и гораздо более прогрессивным рельефом черепа с почти полным отсутствием надбровных дуг, характерных почти для всех ископаемых гоминид (кроме, пожалуй, только неандертальцев группы Схул и кроманьонцев), а также неск. сглаженные костным рельефом. Наиболее молодые находки имеют возраст 1,2 млн. лет.
Моря Сахары и Африки
Не исключено, что этот вид является непосредственным предком палеоантропов группы Схул, хотя между древнейшими людьми сапиентного типа и архантропами лежит достаточно большой отрезок времени: в Эфиопии наиболее древние известны ок. 600 тыс. л. н., в С. Африке (Алжир) — 700 тыс. л. н. (т. н. атлантроп). В то же время не исключено, что потомками рудольфского питекантропа могут быть и карликовые пигмеи — архаичный человеческий тип, аморфный в расовом плане и расселившийся в Ц. Африке от её восточных до западных экваториальных лесов. Можно предположить, что в результате дальнейших межвидовых связей популяций ген этого вида был усилен на позднейшем этапе становления человека, что и привело, в числе многих др. причин, к появлению более грацильного неоантропа со сглаженным костным рельефом.
Пигмеи (греч. Πυγμαῖοι — «люди величиной с кулак») — группа низкорослых народов, обитающих в экваториальных лесах Африки. Другое название афр. пигмеев — негрилли. Совр. пигмеи делятся на западных (бакола, бака, ака в Республике Конго, Камеруне, Габоне и ЦАР) и восточных (мбути на востоке Демократической Республики Конго (ДРК), тва в ДРК, Руанде, Бурунди, Танзании и Уганде).
Рост от 124 до 150 см для взрослых мужчин, кожа светло-коричневая, волосы курчавые, тёмные, губы сравнительно тонкие, этот физический тип можно классифицировать как особую расу. Возможная численность пигмеев может составлять от 40 до 280 тыс. человек. По внешнему виду к ним близки негритосы Ю. Азии и ЮВА, жители нек. о-вов Меланезии и С.-В. Австралии, но генетически между негриллями и негритосами имеются сильные различия.
Для выявления мутаций, объясняющих миниатюрность пигмеев, учёные сопоставили геномы пигмеев из племени батва и их соседей-земледельцев бакига, также для пигмеев-бака и их соседей-земледельцев нзебе и нзими. Выяснилось, что одно-нуклеотидные полиморфизмы, отвечающие за низкий рост, у пигмеев батва и бака отличаются, а значит карликовость у афр. племён является результатом конвергентной эволюции.

Сравнительный рост европейца (слева) и пигмея (справа)


У живущих восточнее бака народов эфе и суа изначально рождаются маленькие дети — ограничитель роста включается во время внутриутробного развития. У бака дети рождаются нормальными, но в первые 2 года жизни дети бака растут заметно медленнее европейцев.
Частота Y-хромосомной гаплогруппы B у пигмеев ака и мбути достигает 35%. В геномах йоруба и пигмеев-мбути обнаружено от 0,2 до 0,7% неандертальских генов. Изучение ДНК биака показало, что примерно 2% генетического материала было вставлено в геном человека ок. 35 тыс. л. н. Эти последовательности они получили от ныне вымершего члена рода Homo, который отделился от совр. человека ок. 700 тыс. л. н.
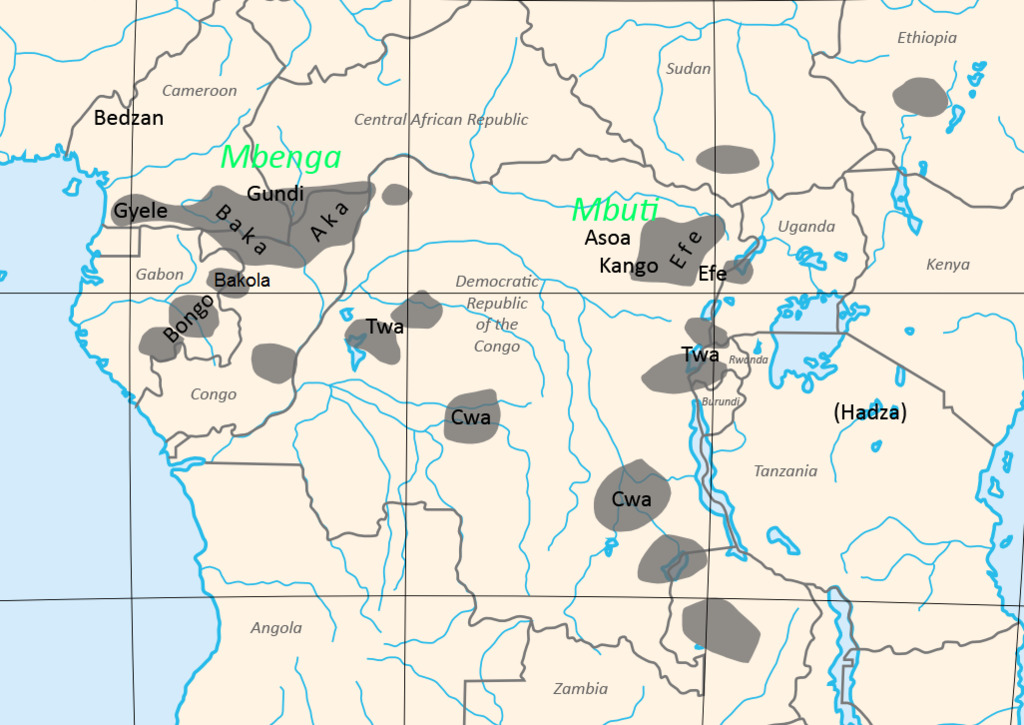
Распространение пигмейских народов согласно Луиджи Кавалли-Сфорца. На карте не отмечены многие народы из группы южных тва
Койсанские народы — условное название группы народов Ю. Африки, представители которых говорят на койсанских языках и/или принадлежат к капоидной расе. Включают бушменов и готтентотов.
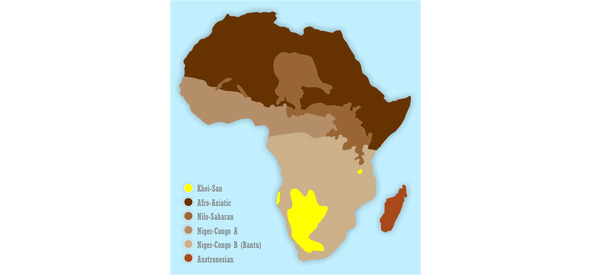

Бушмены
Генетики провели анализ одно-нуклеотидных полиморфизмов (SNP) в хромосомной ДНК представителей разных афр. племён. Согласно опубликованным данным анализа мтДНК, койсанские народы принадлежат к субкладе L0d мт-гаплогруппы L0. Разделение гаплогруппы L0 (мтДНК) на субклады L0k и L0d произошло св. 100 тыс. л. н. в Африке. Обе популяции людей вновь слились в каменном веке, при этом признаки 1-й популяции присутствуют у большинства народов земли, а признаки 2-й популяции наиболее заметны у койсанских народов, живущих на юге Африки. Исследование Y-хромосомы показало, что койсанские народы являются носителями Y-хромосомных гаплогрупп A и B. Совр. койсанские народы имеют до 30% генетического материала, полученного в результате недавнего обмена генами с в.-афр-ми и евразийцами.
Учёными было подтверждено, что койсанские народы с древнейших времён были изолированы от остального человечества. Среди них были обнаружены группы, живущие порознь в течение 30 тыс. лет. Научные работы с результатами исследований вышли в журналах Science и Nature Communications.
Южно-африканская раса (также бушменская, капоидная, койсаноидная) — малая раса в составе большой афр. негроидной, или з.-экваториальной расы, распространённая в пустынных и полупустынных р-нах Ю. Африки. Была предложена как малая раса амер. антропологом Карлтоном Куном в 1962 г. Её представители (бушмены и готтентоты) говорят на языках койсанской ЯС.
Ю.-афр. малая раса довольно сильно отличается как от других негроидов, так и от европеоидов и монголоидов. Сочетание признаков настолько своеобразно, что нек. антропологи выделяют ю.-афр. расу (под названием «капоидная») как 5-ю большую расу наравне с собственно негроидами (негрской и ц.-афр. малыми расами по общей расовой классификации).

В отличие от других негроидов капоиды имеют не чёрный или тёмно-коричневый, а бурый или желтовато-бурый цвет кожи. Волосы и глаза тёмные. Волосы спирально завитые (из-за чего появился миф о слабом росте волос). Нос широкий, с низким переносьем. Третичный волосяной покров слабый. Разрез глаз меньше, чем у негрской малой расы, складка верхнего века развита хорошо, встречается эпикантус. Лицо неск. уплощено, его размеры небольшие. Нижняя челюсть очень грацильна. Черты лица в целом похожи на монголоидные. Длина тела ниже средней. Сильный поясничный лордоз сочетается со значительной стеатопигией (жироотложение на ягодицах), особенно у женщин, но также и у мужчин. У взрослых сильно выражена рано развивающаяся морщинистость кожи.
В составе ю.-афр. расы иногда выделяют 2 антропологических типа — бушменский и готтентотский. Последний отличается более покатым лбом, более развитым надбровным рельефом, высоким переносьем, бо́льшими размерами черепа и бо́льшим ростом.
Капоидов считают остатком какой-то древ. расы южного полушария. Предполагается, что в древности она была распространена более широко, в т. ч. обитала севернее экватора, но была вытеснена из основных р-нов своего расселения — главным образом, негроидами. Особенности этой расы достаточно хорошо фиксируются в ископаемых находках палеолита и мезолита Африки. Представлена у бушменов, готтентотов и значительной части ю.-афр. банту.
Бушменский антропологический тип — один из 2 антропологических вариантов ю.-афр. расы (наряду с готтентотским вариантом). Представлен у койсанских народов бушменской группы (сан), населяющих часть территории Ю. Африки — С.-В. Намибии, Ботсвана, Ю. Ангола, З. Зимбабве и С.-З. ЮАР. Основной особенностью, по которой выделяется бушменский антропологический тип, считают грацильность и меньшие размеры в строении головы и тела в сравнении с более крупным и массивным готтентотским типом.
Бушменский антропологический тип выделяется, в частности, в исследованиях совет. антрополога Г. Ф. Дебеца. Согласно разработанной им классификации рас, бушменский тип вместе с готтентотским образуют бушменскую расу афр. ветви в составе большой негро-австралоидной расы.
Для бушменского антропологического варианта характерны все типичные антроп. признаки ю.-афр. (капоидной, бушменской) расы. Вместе с представителями готтентотского типа бушмены разделяют такие особенности, как распространение более светлых оттенков кожи в сравнении с остальными популяциями негроидов; сильное развитие курчавости волос, зачастую образующих неб. спирально-завитые пучки; слабое развитие третичного волосяного покрова; неб. высота черепа по абсолютным и относительным параметрам в сравнении с высокосводными популяциями прочих негроидов; узкий нос с очень плоским переносьем; своеобразный рисунок ушной раковины; сильно выраженный поясничный лордоз; относительно неб. длина тела; напоминающие монголоидные признаки сильная уплощённость лица и распространение эпикантуса; частые случаи стеатопигии и т. д.
В сравнении с готтентотским антропологическим типом бушменский тип характеризуется такими особенностями, как:
меньшая покатость лба,
меньшее развитие надбровных дуг,
более низкое переносье,
меньшие размеры черепа в целом, в особенности меньшая высота черепа,
значительно менее высокий рост.
Особое место в антропологическом плане занимают исчезнувшие к наст. времени береговые бушмены, которые некогда населяли побережье на ю.-з. Африки от севера Намибии до Кейптауна. Их антропологические признаки, в частности, размеры черепа, имели промежуточные значения между значениями признаков бушменского и готтентотского типов. Более того, береговые бушмены отличаются и от бушменов, и от готтентотов наличием мезокранных и даже брахикранных черепов, для них также характерны наиболее короткое основание черепа, низкие глазницы и наименьшее выступание вперёд челюстей.
Этноязыковая принадлежность бушменов и их антропологические особенности не всегда совпадают. Нек. представители бушменского типа говорят, например, на диалектах готтентотского языка нама, а у части готтентотов, в частности, у разных групп нама, обнаруживается бо́льшая генетическая близость к соседним группам бушменов, нежели к остальным готтентотам. Многие группы бушменов в разное время были ассимилированы соседними с ними народами, относящимися к негрской расе, что привело к проявлению нек. бушменских признаков в антропологическом облике бантуязычного населения Ю. Африки.
Готтентотский антропологический тип — один из 2 антропологических вариантов ю.-афр. расы (наряду с бушменским вариантом). Представителями его являются койсанские народы готтентотской группы (кой-коин), живущие в настоящее время в Ю. и Ц. Намибии, а также отчасти в Ботсване и в ЮАР. Готтентотский антропологический тип традиционно рассматривается как более крупный и массивный в сравнении с более мелким и грацильным бушменским типом.
Готтентотский антропологический тип выделяется, в частности, в работах совет. антрополога Г. Ф. Дебеца. В его классификации человеческих рас готтентотский тип вместе с бушменским образуют бушменскую расу афр. ветви в составе большой негро-австралоидной расы.
Вместе с бушменским антропологическим типом готтентотский тип разделяет все основные антроп. признаки ю.-афр. (капоидной, бушменской) расы. К таким признакам относятся: более светлый цвет кожи в сравнении с остальными афр. негроидами (часто жёлто-бурого оттенка); более развитая, чем у прочих негроидов, курчавость волос (волосы чаще всего спирально-завитые, спутанные в неб. пучки); слабое развитие третичного волосяного покрова; абсолютно и относительно низкая высота черепа в отличие от высокого свода черепа у большинства негроидов; узкий нос с очень плоским переносьем; сравнительно низкий рост; высокая степень уплощённости лица; большая частотность распространения эпикантуса; нередко отмечаемая стеатопигия и другие признаки.
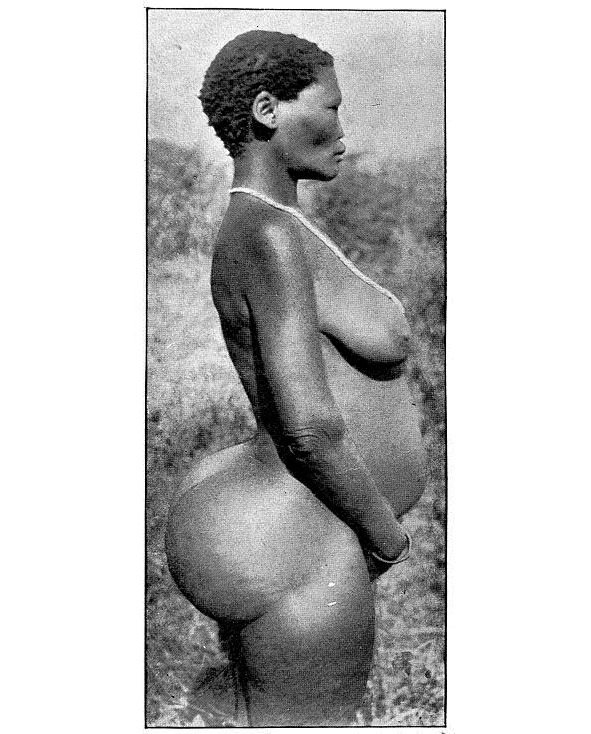
В сравнении с бушменским антропологическим типом готтентотский тип характеризуется такими особенностями, как:
бо́льшая покатость лба,
большее развитие надбровных дуг,
менее низкое переносье,
бо́льшие размеры черепа в целом, в особенности бо́льшая высота черепа,
значительно более высокий рост.
Помимо собственно готтентотских народов часть признаков готтентотского типа отмечается у метисной группы рехоботских бастардов (бастеров), потомков готтентотов и голландских колонистов, живущих в Намибии. Нек. народы, говорящие на готтентотских языках, не принадлежат к готтентотскому антропологическому типу. Например, дамара, говорящие на языке нама — у них отмечаются признаки негрской расы. О том, что этноязыковая принадлежность не всегда коррелирует с происхождением и антропологическим типом, говорят новейшие генетические исследования. Согласно этим исследованиям, нек. группы готтентотов оказываются генетически ближе к бушменам, нежели к остальным готтентотам, например, готтентоты-нама.


Представители готтентотских народов

Представители готтентотского народа нама (нач. 20 в.)
По предположениям нек. антропологов, отличия признаков готтентотского типа от бушменского возникли в результате частичной метисации бушменов с представителями негрской расы. Также на изменение облика готтентотов, возможно, повлияла смена образа жизни и типа питания при переходе от собирательства к скотоводству.
Промежуточной в расовом плане формой являются и совр. полинезийцы, сочетающие в своём типе черты европеоидов (ю.-амер. происхождения), монголоидов (австронезийского происхождения), американоидов и даже австралоидов (из Австронезии).
Уже ок. 2 млн. л. н. в Европе (и Якутии в Азии) появляются чопперы, относящиеся к Олдувайской культуре.

Фильм: https://my.mail.ru/v/pravilnyperevodfilmov/video/18/580.html
Формация Homo erectus (архантропов) уже отличалась тем, что умела создавать из камня примитивные скребки, свёрла, строить хижины, покрытые шкурами, пользоваться огнём (с определённого периода, чем не владел ближайший родич Homo sapiens — водный вид, русалка, что стало первым отличием древнейшего человека от водного вида), охотиться (главное оружие — копьё, для твёрдости обожжённое на огне) и ловить рыбу. Мясо было главной пищей (в отличие от рыбы — главной пищи водного вида; что позволяет сделать вывод о том, что охота была освоена прото-человеком гораздо позже рыбной ловли). Нек. пороки костей Homo erectus говорят о неумеренном потреблении костного мозга и сырой печени, о переизбытке витамина А. Отмечается, что у афр. архантропов объём мозга был меньше, чем у европ. и азиатских (что, видимо, связано, в известной мере, со средой обитания).
Находки Homo heidelbergensis
Уже ок. 780—750 тыс. л. н. в Европе (очаги на юге Франции) использовался огонь, не менее 500 тыс. л. н. — в В. Азии (Китае). Ок. 800 тыс. л. н. в Европе уже отмечался вид гейдельбергского питекантропа, сближающегося по многим позициям с атлантропом мавританским, населявшим С. Африку (Алжир), и более прогрессивного по сравнению с более древ. «классическими» питекантропами (с.-афр. архантроп, морфологически сходный с Homo erectus, возраст большинства находок — 360 тыс. лет, впервые обнаружен в 1954—1955 гг.; название atlanthrop (в честь Атласских гор С. Африки) часто используется без ссылки на конкретный вид).

В то же время для этих видов отмечается большое сходство с синантропом. Связь гейдельбергского питекантропа с атлантропом, и появление его в Европе неудивительны, поскольку в то время ещё существовал Сицилийский перешеек, хотя миграции могли идти и из М. Азии через Балканский п-ов. В то де время, как будто, в Европе отмечается неск. отличный тип архантропа — т. н. «прогрессивного». Впрочем, это может быть и один вид, но последний, вероятно, более поздний, и непосредственно предшествующий палеоантропам. Он отмечен в Венгрии и В. Германии. Представители архаического вида Homo sapiens появляются в Англии и Германии ок. 500 тыс. л. н. — т. н. пре-сапиенсы, давшие начало местным палеоантропам.
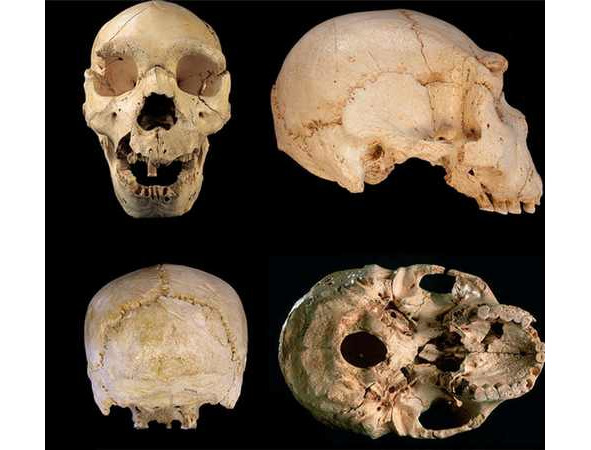
Череп гейдельбергского человека
Т. о. Европу населили архантропы прото-европеоидной расы, восходящей к Y-хромосомной гаплогруппе С (ныне превалирующей у типичных с.-монголоидов Ц. Азии) — которым для того, чтобы утратить избыточную пигментацию, был необходим весьма длительный период, исчислявшийся сотнями тыс. лет в условиях существования в холодной приледниковой зоне. При этом, если предположить, что предки айнов австралоидного расового ствола населили ДВ ок.100 тыс. л. н. и полностью не утратили пигментацию, следовательно, этого времени не достаточно для процесса депигментации. Тот же процесс проходили и предки монголоидов, расселившиеся в Ц. Азии (например, в высокогорном Тибете многие монголоиды имеют тёмную кожу, ничем не отличающуюся от капоидной или негроидной).

Череп Homo heidelbergensis
Ок. 300 тыс. л. н. на Яве появляется т. н. солоский питекантроп или яванский неандерталец, именуемый также нгандонгским человеком, считающийся поздней формой яванского питекантропа, по объёму мозга близкий к синантропу и имеющий примитивные особенности в строении черепа, и гораздо более близкий к питекантропам и синантропам, чем к палеоантропам. По выражению Бьёрна Куртена, нгандонгский человек был «поразительно примитивным, недалеко ушедшим от Homo erectus, хотя жил в верхнем плейстоцене». Впрочем, не исключено, что нгандонгский человек — предок, наоборот, папуасов или негритоссов, а родственный ему афр. тип, соответственно — нагроидов. Однако, в т. сл. они должны были уже оттеснить австралоидов на юг, но те появляются в Австралии как будто не ранее 50 тыс. л. н. (для Новой Каледонии, где обитает отдельный австралоидный новокаледонский тип, период заселения не известен). В то же время существует также «тасманийская загадка», поскольку неизвестно, когда тасманийцы, относящиеся к негроидному стволу, попали в Тасманию.
Родственный яванскому неандертальцу тип обитал в Африке, южнее Сахары, однако «исчез» 30 тыс. л. н. Отмечено, что сходство нгандонгского человека с з.-афр. неандертальцем не абсолютное: в строении его черепа обнаруживается целый ряд примитивных особенностей: очень сильно развитый саггитальный валик (идущий вдоль всей черепной коробки по её средней линии), необычайное разрастание рельефа черепа. По признаниям исследователей, он едва ли не синхронен палеоантропам, а быть может, даже древнейшим представителям совр. людей. Возможно также, что вид, родственный нгандонгскому неандертальцу, расселявшийся в З. Африке, преемствует атлантропу, сближающемуся с гейдельбергским питекантропом. Но именно в з.-афр. р-нах живут совр. фульбе австралоидной ветви (а в нагорье Тибести — тиббу, именуемые Геродотом «пещерными эфиопами», которых нек. антропологи пытаются относить даже к европеоидам), которые часто темно-пигментированы, что обычно считается негроидной примесью, но именно это позволяет заключить, что они никогда не жили в северных условиях. Но антропологически они действительно близки европеоидам — как предки (видимо, носители Y-хромосомной гаплогруппы Е). В данном случае уже можно говорить об очень древ. расовом родстве.
В 1936 г. в карьере близ Моджокерто (на о. Ява) был найден детский череп Pithecanthropus V. Также в 1930-е гг. в бассейне р. Соло на Яве (в т. ч. возле Сангирана и у расположенного недалеко оттуда пос. Нгандонг) были раскопаны дополнительные кости гоминида, чертами сходного с питекантропом, но более молодого: возраст самых поздних костей составляет менее 100 тыс. лет. Первоначально этот гоминид был описан как Homo (Javanthropus) soloensis. Явантропа из Нгандонга при схожем с яванским питекантропом архаичном строении черепа, указывающем на преемственность, отличает неск. больший объём мозга — в среднем ок. 1150 куб. см. Соответственно, многими учёными явантропы в 21 в., подобно синантропам, рассматриваются как представители вида Homo erectus. Другие авторы продолжают рассматривать их как отдельный вид Homo soloensis, либо выделяют наиболее молодые фоссилии в качестве костей раннего сапиенса — Homo sapiens soloensis.

Явантроп (нгандонгский, или солоский человек, лат. Homo erectus soloensis) — название ископаемого человека из Индонезии. Кости явантропов были обнаружены в 1931—1933 гг. на левом берегу р. Соло, у дер. Нгандонг (о. Ява), среди них — 12 неполных мужских и женских черепных коробок. Изначально был классифицирован как Homo sapiens soloensis, ныне классифицируется Homo heidelbergensis или Homo erectus. Последние Homo erectus из Нгандонга жили в период 117—108 тыс. л. н.
Антропологов давно волнует вопрос появления синантропа в В. Азии. К.б.н, доцент кафедры антропологии биологического факультета МГУ им. Ломоносова, С. В. Дробышевский, например, безапелляционно утверждает так: «Монголоиды появляются в Азии значительно позже! Это 10—15 тысяч лет назад. Даже Чжоукоудянь (верхняя пещера) — там все 3 черепа сапиенсов — не монголоидные. Американские индейцы — сами по себе живой пример, что монголоиды сложились поздно, т. к. индейцы — это „прото-монголоидный“ вариант. А заселение Америки — как бы его не оценивали — это 15 тыс. л. н., а скорее всего даже меньше. Т.е. 15 тысяч лет назад монголоидов или не было, или они были очень маленькой группой».
https://antropogenez.ru/interview/194/
Т. е. он не видит никакой связи с капоидной расой Ю.-З. Африки, и сходства синантропа по многим позициям с нын. монголоидной расой :) Это, конечно, опрометчиво.
Наиболее древ. представители пекинского питекантропа (синантропа) известны ок. 400 тыс. л. н., но, поскольку огонь здесь уже использовался на 100 тыс. лет ранее, можно полагать, что появление именно предков синантропа положило начало распространению здесь человека. Объём мозга синантропа варьировал в пределах 850—1220 куб. см (по др. данным, 915—1225 куб. см). Для синантропа характерны явные монголоидные признаки. Наиболее поздние его представители отмечаются до 220 тыс. л. н., т. е. примерно до того периода, когда с истор. арены исчезает Homo erectus. Т. о., говоря о происхождении монголоидов, надо отметить, что в среднем палеолите (мустье) их расселение отмечено в С. и Ц. Китае, т. е. в В. Азии, по меньшей мере, с периода 500—400 тыс. л. н. — с расселения синантропа. Однако нам неизвестна зона их расселения в нижнем палеолите, хотя можем предполагать, что это П., Ср. и Ц. Азия. Сходство в нек. плане гейдельбергского питекантропа и синантропа — только внешнее, не имеющее близко-родственных корней. Гораздо больше сходства между совр. монголоидами и америндами (особенно северной ветви), восходящими к алтайскому/денисовскому архантропу/палеоантропу.
Исходя из вышесказанного, следует, что синантроп не мог прийти в В. Азию из Ю. Азии или ЮВА, и уж тем более не мог расселяться из Африки через Европу (появление негро-папуасов в Ю. Азии и ЮВА относится-де к периоду 40—35 тыс. л. н., а монголоидов — ещё гораздо позднее — в позднем неолите). Т. о. отношений прямого характера, равно как и близкого родства, между синантропом и гейдельбергским человеком быть не могло. Следовательно, двигался он с Б. Востока (100 тыс. л. н. в Леванте как будто отмечены палеоантропы с монголоидными чертами, однако, даты не абсолютны и это может быть фиксацией лишь реликтовой остаточной популяции).
Алтайский/денисовский архантроп/палеоантроп (Homo altaensis), возрастом 170—30 тыс. л. н., является предком америндского ствола, связанного с Y-хромосомной гаплогруппой Q, в значительном количестве характерной для енисейских кетов (95%), обских селькупов (70%), туркмен (до 50%), приуральских угров — хантов и манси (ок. 25%). Его потомки в своей массе мигрировали на Американские континенты, как считается, в период ок. 15—10 тыс. л. н. (а, по всей вероятности, намного раньше) через гипотетическую Берингию — якобы существовавший сухопутный мост между Чукоткой и Аляской. Учитывая происхождение, можно предположить, что носители этого типа освоили степные пространства к востоку и мигрировали вдоль горно-лесной зоны Хангая, Саяна и р. Амур, через о. Сахалин, о-ва Курильской гряды и Камчатский п-ов.

Портрет денисовской девочки, основанный на предсказанных особенностях строения черепа
Уже ок. 300 тыс. л. н. в Китае появляется вид т. н. ляонинского человека, череп которого имел нек. черты сходства с Homo erectus: низкий лоб, выступающие вперёд надбровные дуги, большие коренные зубы. Однако в черепе заметны и признаки строения черепов Homo sapiens: округлой формы черепная коробка, больший объём мозга — до 1390 куб. см, тонкие стенки затылка. Морфология «ляонинца» была более прогрессивной по сравнению с синантропом. Последние, наиболее молодые, находки датируются 125 тыс. л. н., что позволяет предположить о том, что этот вид является предком америндов. По-видимому, он является прямым предком Homo longi («человек-дракон», из Харбина), возрастом ок. 150 тыс. лет. Как и у Homo sapiens, у Homo longi был большой мозг, расположенный на относительно коротком лице и неб. скулах. Но такие черты, как толстые надбровные дуги, большие коренные зубы и почти квадратные глазницы, напоминают неск. вымерших популяций или видов Homo, включая Homo heidelbergensis и неандертальцев. Эти виды относятся к ключевому периоду эволюции человека — среднему плейстоцену, который длился от 789 000 до 130 000 л. н. При этом он удивительно похож на «денисовца», и резко отличается от монголоидов.

Так, предположительно, выглядел Homo longi. Как говорится, найдите 10 различий

Так, вероятно, Homo longi смотрелся в лесной обстановке

Ископаемые человеческие черепа из Китая. Home erectus (пекинский человек) находится слева, а примеры недавно определенной линии Homo longi — справа. Череп Харбина — последний справа
Филогенетическое древо — дерево, отражающее эволюционные взаимосвязи между различными видами или другими сущностями, имеющими общего предка
Используя статистические сравнения с другими окаменелостями эпохи среднего плейстоцена из Африки, Азии и Европы, исследователи пришли к выводу, что у Homo longi и Homo sapiens был общий предок, который жил около 949 тыс. л. н. Считается, что общий предок неандертальцев и совр. людей (Homo sapiens) жил на Земле чуть более 1 млн. л. н. Если это так, то родство Homo longi с Homo Sapiens намного больше, чем между нами и неандертальцами. Исследователи также полагают, что Homo longi был современником эволюционирующих азиатских ветвей Homo sapiens — неандертальцев и денисовских людей. При этом стоит отметить богатое видовое многообразие в одну и ту же эпоху Азиатском регионе: западным соседом его были синантроп — предок монголоидов, и алтайский/денисовский палеоантроп — предок америндов.
Амуд I часто расценивается как классический неандерталец, но по целому ряду признаков, особенно лицевого скелета, оказывается намного сапиентнее, чем гоминиды из пещер Схул и Кафзех. Например, относительные размеры верхней челюсти значительно меньше, чем у неандертальцев Европы, а форма альвеолярной дуги не отличается от совр/, хотя альвеолярный отросток спереди заметно уплощён. Сапиентны форма альвеолярной дуги и соотношение мыщелковой и угловой ширины нижней челюсти. В отличие от схульцев, таз Амуд I сравнительно небольшой, с довольно короткой лобковой костью.

Череп Амуд I (55—45 тыс. л. н.)

Череп Схул V
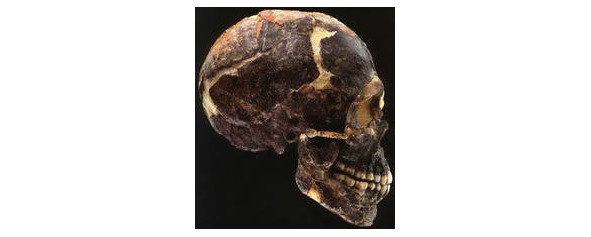
Череп Кафзех 9
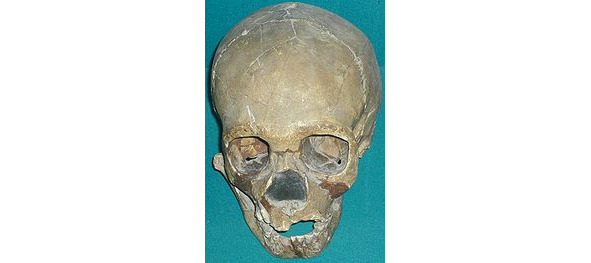
Череп из Тешик-Таш
Кебара II (последний справа) для сравнения с Табун (слева) и схул-кафзехцами
Алтайский/денисовский архантроп/палеоантроп отчасти сходен с мезолитическим расовым типом Ю. Урала, восходящим к верхне-палеолитическому населению П. Азии (в т. ч. Ирака и Ирана), где в среднем палеолите жил т. н. передне-азиатский/палестинский палеоантроп/неандерталец (от М. Азии до Междуречья и Ср. Азии; Израиль: пещеры Амуд, Кебара, эс-Схул и эт-Табун, Ирак: пещера Шанидар, Ср. Азия: пещера Тешик-Таш), известный с 70 тыс. л. н. (т. е. тогда, когда по ряду данных, происходит исчезновение из Европы классических неандертальцев), имеющий многие примитивные черты (их отличает, как и африканский тип, необычайная массивность лицевого скелета, а также значительно развитый рельеф черепа) в сочетании с нек. прогрессивными признаками (высокий свод черепа, почти прямой лоб, более четко очерченный подбородочный выступ, менее выраженный надглазничный рельеф, удлинённые верхние и нижние конечности, высокий рост, пропорции, характерные для совр. людей), и относившиеся к Y-хромосомной гаплогруппе G1, исходное происхождение которых восходит к Y-хромосомной гаплогруппе С. Потомки этого типа в мезолите населили Ю. Урал, мигрировав из Ю. Прикаспия.
Утверждается, что передне-азиатские палеоантропы — это неандертальцы и неандерталоиды такие как Кебара, Табун, Амуд, Шанидар и Схул-Кафзех. Однако, Схул (135—100 тыс. л. н.), Кафзех (99—90 тыс. л. н.), Амуд (55 тыс. л. н.) и Табун — безусловно особые неандертало-сапиентные люди. Но они рубеж в 65—55 тыс. л. н. не перешагнули, а с архаичным неандертальцем ср.-аз. пещеры Тешик-Таш их отношения не ясны. А вот Шанидар I — обычный, более того, поздний (51—45 тыс. л. н.) неандерталец, как и классический неандерталец Кебара II (60—48 тыс. л. н.). Скорее всего, оба — мигранты-вытеснители схул-кафзехцев из Европы.
Т. о. переднеазиатские палеоантропы являются всего лишь дальними боковыми родичами европейского классического неандертальца.
Передне-азиатским палеоантропам отчасти близки европейские атипичные неандертальцы, жившие во время Рисс-Вюрмского межледниковья (150—70 тыс. л. н.), которые характеризовались сочетанием архаических и прогрессивных черт в строении черепа, объем которого был меньше, чем у классических форм (находки из Эрингсдорфа (Германия), Саккопасторе (Италия), Гановце (Словакия), Крапина (Хорватия) и др.); в наст. время их относят к виду гейдельбергского человека.
Появление 700—500 тыс. л. н. новой формации — палеоантропов, зачастую имевших самое мощное физическое строение и самый большой мозг, возникшие как ответ на суровые условия природной среды — неоднородно повлияло на географию и численный состав населения. Периодом 290—140 тыс. л. н. датируется появление предтечи человека совр. вида — неоантропа, известного как пре-сапиенс. С палеоантропами связана средне-каменная) культура Мустье, выходящая по сравнению с нижним палеолитом (Шелль, Ашель) на более высокий качественный уровень, характерный началом формирования собственно культуры (с возникновением 1-х религиозных представлений).
Выяснилось, что у неандертальцев:
— была светлая кожа,
— волосы оттенков блонд, светло-русые или рыжие,
— голубые, серые, светло-зеленые глаза,
— гиперлордоз.
Периодом 250—110 тыс. л. н. датируется существование в Европе прогрессивного/атипичного палеоантропа/неандертальца, которого будто бы 110 тыс. л. н. сменяет классический палеоантроп/неандерталец. У 1-х были черты, близкие как архантропам, так и неоантропам. 2-е были более примитивны по ряду позиций, но очень чётко приспособлены к суровому климату. Впрочем, ряд исследователей (например, член-корр. В. П. Алексеев), указывают на сходный ареал обитания и отмечают, что представители 1-го типа представлены в своей основе женскими, а представители 2-го — преимущественно мужскими черепами. 1-х относят к группе Эрингсдорф, 2-х — к группе Шапёлль-Ферасси. Различия между «классическими» и «атипичными» палеоантропами Европы довольно резкие: мозг более ранних, правда, меньше по объёму, но более прогрессивен по строению — череп выше, рельеф черепа меньше (исключение составляют сосцевидные отростки, развитые сильнее — типично человеческий признак), на нижней челюсти намечается подбородочный треугольник, размеры лицевого скелета меньше. С ними сходны с.-афр. находки из Джебель-Ирхуда и один из черепов, найденный при раскопках пещеры Схул в Палестине/Израиле, известный как Схул-9 (наиболее примитивный из группы Схул).
135 тыс. л. н. неандертальцы распространились не только в Европе, но и в Азии и С. Африке, хотя физически отличались в более слабую сторону от своего классического европ. современника. Ок. 120 тыс. л. н. (межледниковый период) неандертальцы освоили Б. Восток и Ср. Азию. Позднее других палеоантропов отмечается в.-азиатский (теперь его место занял Homo longi).
Европейские неандертальцы населяли также Крым, запад М. Азии и С. Африку.